- Original Article
- Published:
Symbiotic association between Solanderia secunda (Cnidaria, Hydrozoa, Solanderiidae) and Medioantenna variopinta sp. nov. (Annelida, Polychaeta, Polynoidae) from North Sulawesi (Indonesia)
Helgoland Marine Research volume 65, pages 495–511 (2011)
Abstract
A mimic scale-worm was found associated with the athecate hydroid Solanderia secunda, commonly found on reefs of the NW coast of North Sulawesi, Indonesia. The species resembled Medioantenna clavata Imajima 1997, which was originally described without any reference to a symbiotic mode of life and later reported to be living on a solanderiid hydroid both in Japanese waters. A detailed morphological analysis led us to consider the Indonesian specimens as a new species, namely Medioantenna variopinta sp. nov., which is congeneric with the Japanese species. The new species differs from the type material of M. clavata as it has elytra with one prominent finger-like papilla and all neurochaetae with unidentate tip, instead of an elytral lump and both unidentate and bidentate neurochaetae on segment two. In turn, the Japanese worms associated with Solanderia are here referred to our new species. Two morphological features in M. variopinta sp. nov. are rather unusual among scale-worms. One of them is its extremely high level of bilateral asymmetry and antero-posterior variability in elytral distribution and the other one is its elongated, upwardly directed nephridial papillae. The morphology and geographical distribution of the host together with the known characteristics of the symbiotic association have also been highlighted.
Introduction
The Class Hydrozoa includes a large variety of life forms that are often involved in symbiotic relationships with other invertebrates (Boero and Bouillon 2005; Gili and Hughes 1995; Puce et al. 2008). Their rather rich symbiotic fauna includes polychaetes (Genzano and San Martín 2002; Lagardère and Tardy 1980), and some species of Teissiera or Hydractinia tend to be specifically associated with polychaetes (Bouillon and Boero 1987; Bouillon 1994; Galea et al. 2007).
Most polychaetes associated with hydrozoans belong to the families Polynoidae and Syllidae, but a few occur among the Lumbrineridae, Spionidae and Serpulidae (Martin and Britayev 1998). The symbiotic syllids show many different relationships with their hydrozoan hosts, from temporal feeding visitors (i.e., predators) to permanent, specific symbionts (Britayev and San Martín 2001; Gidholm 1966; Nygren 2004). In turn, the polynoids (or scale-worms) associated with hydrozoans are in general more diversely and closely related with their hosts. Scale-worms are predominately found in association with stylasterid or antipatharian corals (Martin and Britayev 1998). They can modify the host colony growth causing the formation of galls with open tunnels (i.e., the so-called worm runs) that facilitate movement of the worms (Britayev 1981; Cairns 1987; Hanley and Burke 1991; Molodtsova and Budaeva 2007; Pettibone 1989, 1991).
In turn, the associations between scale-worms and other hydrozoans such as the hydroidomedusae seem to be scarce, with only two known records: Lagisca zibrowii Hartmann-Schröder 1992, associated with Pseudosolanderia sp. in relatively deep waters off New Caledonia (Hartmann-Schröder 1992), and Medioantenna clavata Imajima 1997, found on a solanderiid colony in the shallow waters off Ogasawara Islands (Japan) (Nishi and Tachikawa 1999). In the latter, however, the hydroid identification looked doubtful. It was reported as “Solanderia misakiensis”, a mis-spelling of S. misakinensis (Inaba 1892), in the abstract and then referred to as S. secunda (Inaba 1892) within the polychaete description. Regardless of these name problems, that paper represents the first reference to a symbiotic mode of life for Medioantenna. Originally, the genus (and its single species known to date, M. clavata) was described from Sagami Bay without any reference to its mode of life (Imajima 1997).
As such, our report on the specimens from North Sulawesi (Indonesia) is the third for the genus. These worms fit well with Nishi and Tachikawa (1999)’s description. However, they clearly differ from the original description of Medioantenna clavata by Imajima (1997), as confirmed by checking an available paratype. Therefore, here, we are describing them as a new species within the genus, namely M. variopinta sp. nov., which includes specimens from Indonesia and from Ogasawara Islands. We have also highlighted the morphological characteristics and geographical distribution of the host, together with the known characteristics of the symbiotic association.
Materials and methods
The presence of a symbiotic polychaete exhibiting well-defined color mimicry was observed on colonies of the Anthomedusan hydroid Solanderia secunda from North Sulawesi (Indonesia), an area that is considered to be a hot spot of marine biodiversity together with the Philippines and Papua New Guinea (Hoeksema 2007).
The taxonomy and ecology of the hydroids of this area have been studied since 2002, when several colonies of the two solanderiid species—Solanderia secunda and S. procumbens—were analyzed both underwater and in the laboratory. However, the first symbiotic polychaetes were obtained between 2005 and 2008, after randomly samplings of the solanderiid colonies by SCUBA diving at different sites of the Bunaken Marine Park and at Mapia House Reef, at a depth ranging between 10 and 30 m (Fig. 1). The scale-worms were always found on S. secunda colonies and never on S. procumbens.

Map of the studied area in the Indonesian archipelago. Sampling sites: 1. Siladen Onong; 2. Siladen Jeti; 3. Tanjung Pisok; 4. Mapia House Reef; 5. Bunaken Timur; 6. Negeri. Map source: http://www.gebco.net/
During February and July 2008, additional diving surveys were conducted to check the infestation of S. secunda colonies at Siladen Onong and Bunaken Timur. At these sites, horizontal transects of about 100 m length were carried out at three depths (10, 15 and 30 m). The occurrence and number of worms in each colony was noted (Table 1). Any attempts to collect the symbionts in situ resulted either in an escape behavior (as the worms tried to hide themselves within the basal portion of the colony) or autotomising. To obtain entire specimens, fragments of the host hydroid were cut with surgical scissors and kept in plastic bags together with the worms. In the laboratory, the hydroid branches hosting polychaetes were observed in vivo under a stereomicroscope to understand the peculiarities of the behavior of the symbiont. Later, some polychaete specimens were carefully separated from the host and preserved either in 4% neutralized formaldehyde or in 95% ethanol.
Both the symbiotic polychaetes and host colonies were examined by scanning electron microscopy (SEM). The target fragments were carefully rinsed with distilled water, dehydrated in a graded ethanol series, critical point dried and coated with gold–palladium in a Balzer Union evaporator. They were observed under a Philips XL20 microscope.
Relevant morphological features were drawn with the help of a camera lucida; then, drawings were scanned and digitized with a Wacom tablet. Body length (L) was measured from the anterior prostomial margin to the posterior border of the last segment. Body width was measured at the level of the widest segment, parapodia excluded (WW) or included but excluding (WP) or non-excluding chaetae (WC). In the “Material examined” and “Measurements” sections, ‘cs’ means complete specimens and ‘mf’ refers to mid-body fragments. The types of the new species are deposited in the “Museo Nacional de Ciencias Naturales” (MNCN) of Madrid (Spain). The types of Medioantenna clavata were loaned by the National Science Museum of Tokyo (NSMT).
Results
-
Phylum ANNELIDA
-
Class POLYCHAETA
-
Family POLYNOIDAE Kinberg 1856
-
Medioantenna Imajima 1997, emended
-
Type species. Medioantenna clavata, Imajima 1997
Diagnosis
Body dorso-ventrally flattened, long, with up to 83 chaetigers and up to 38 pairs of elytra. Elytra on segments 2, 4, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 26, 29, 32, 35 and thereafter with attachment varying between specimens and between left and right side of the same specimen, to end of body, with lump or large finger-like papillae on rear margin. Dorsum more or less covered by elytra. Body with dorsal tubercles on cirrigerous segments. Prostomium bilobed, with small cephalic peaks and three antennae. Lateral antennae small, inserted ventrally to median antenna. Median antenna with large ceratophore and style, inserted antero-dorsally on prostomium. Two pairs of eyes and two palps present. Tentaculophores achaetous. Proboscis with nine pairs of papillae and two pairs of jaws. Parapodia relatively long, biramous; both noto- and neuropodia with elongate acicular lobes; notopodia, small, conical, located at antero-dorsal side of neuropodia; neuropodia with a supra-acicular process; tips of noto- and neuroacicula penetrating epidermis. Notochaetae slightly stouter than neurochaetae, straight to falcate with slight serrations and unidentate tips. Neurochaetae more numerous, with strong serration and unidentate or bidentate tips. Median antenna, tentacular and dorsal cirri with subdistally inflated styles and filiform, pointed tips.
Remarks
Imajima (1997) placed Medioantenna in the Arctonoinae. However, the position and size of median antenna and the very small spur-like lateral antennae of the genus are very unusual features for this subfamily. Moreover, neither Imajima (1997) nor Nishi and Tachikawa (1999) mentioned the presence of dorsal tubercles that occur on the tentacular segments in M. variopinta sp. nov. and are clearly visible in Imajima’s drawings (Imajima 1997; Fig. 8b, c). The presence of dorsal tubercles also disagrees with the subfamily diagnosis. As the separation of Arctonoinae from Polynoinae is still not clear enough, we agree with Barnich and Fiege (2001, 2003) that a revision of the genera should be included within the former. Therefore, we prefer to avoid the inclusion of the genus in any subfamily of polynoids.
Medioantenna was established by Imajima (1997) for Medioantenna clavata Imajima, 1997 and has been considered as monotypic to date. Our findings confirmed the existence of a second species of the genus, M. variopinta sp. nov., which is found on tropical hydroids in the coastal waters of Japan and North Sulawesi and which is fully described here below.
Examined material
Paratype: NSMT-Pol R74, two fragments. Kannonzukadashi, Amadaiba, 80 m deep, March 20, 1951 according to Imajima (1997) (label written in Japanese).
Remarks
The morphological characteristics of the paratype fully agree with the original description by Imajima (1997). However, the following details were not previously mentioned, and as such, have been reported here to complete the description of the species. In the observed paratype, the number of notochaetae per bundle ranged from 13 to 16 (on chaetigers 2, 12, 23 and 39). The nephridial papillae become elongated, directed upward between parapodia to reach one-third of the parapodial height from chaetiger 3 to posterior end. The paratype was thus a female measuring L: 25 mm (20 + 5 mm), WW: 2.9 mm, WP: 6 mm, WC: 10.2 mm, 35 (30 + 5) segments.
-
Medioantenna variopinta sp. nov. (Figs. 2, 3, 4, 5, 6, 7, 8, 9)
Fig. 2 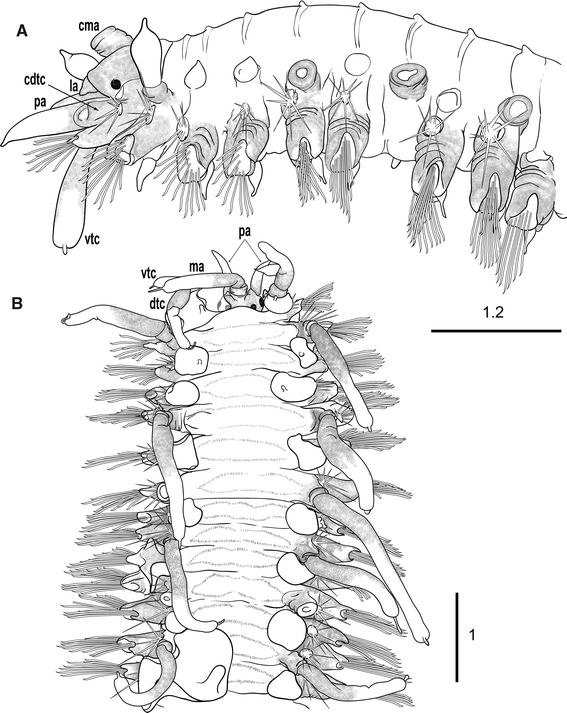
Medioantenna variopinta sp. nov. Holotype. a. Anterior end, lateral view. b. Anterior end, dorsal view; ma—median antenna, cma—ceratophore of median antenna, cdtc—cirrophore of dorsal tentacular cirrus, dtc—dorsal tentacular cirrus, vtc—ventral tentacular cirrus, la—lateral antenna, pa—palp. Scale bars are in mm
Fig. 3 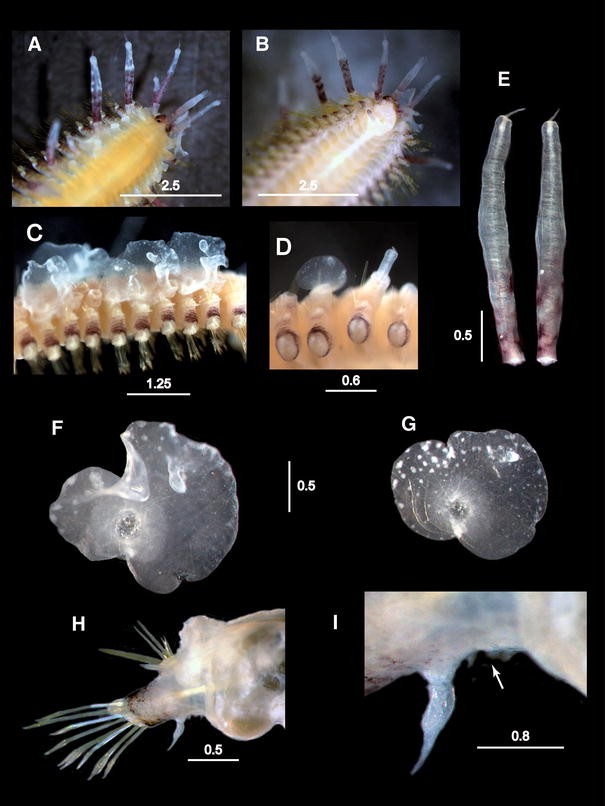
Medioantenna variopinta sp. nov. Preserved specimen. a. Anterior end, dorsal view. b. Anterior end, ventral view. c. Mid-anterior region with elytra attached (anterior end toward the left). d. Mid-posterior region with one regenerating elytron and dorsal cirri (anterior end toward the left). e. Detached dorsal cirri from mid-body. f. Mid-anterior elytron. g. Mid-posterior elytron. h. Mid-body elytrophorous parapodium in postero-anterior view. i. Detail of the ventral side of the same parapodium, showing the ciliated papillae (white arrow). Scale bars are in mm
Fig. 4 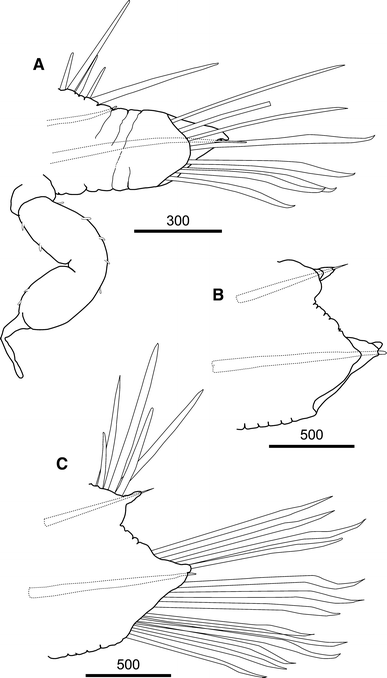
Medioantenna variopinta sp. nov. Holotype. a. Parapodium from the second segment, antero-posterior view, showing the long ventral cirrus. b. Parapodium from mid-body (39 segment), antero-posterior view. c. Same as b, postero-anterior view. Scale bars are in μm
Fig. 5 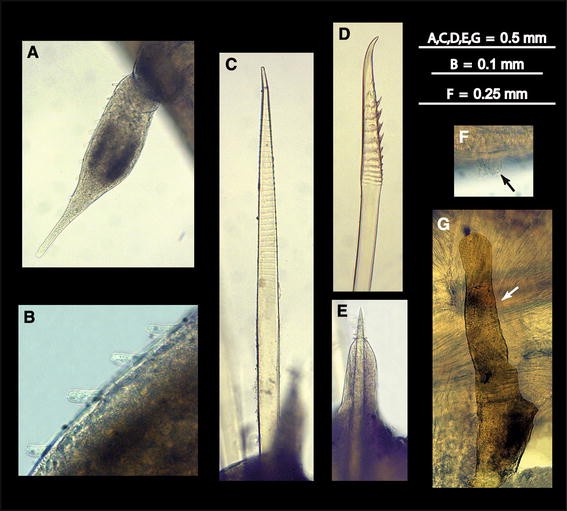
Medioantenna variopinta sp. nov. Preserved specimen. a. Ventral cirrus. b. Detail of the papillae from the same. c. Notochaeta. d. Neurochaeta. e. Notoacicula and notoacicular lobe. f. Cilia from the ventral ciliated mounds (black arrow). g. Nephridial papilla (white arrow)
Fig. 6 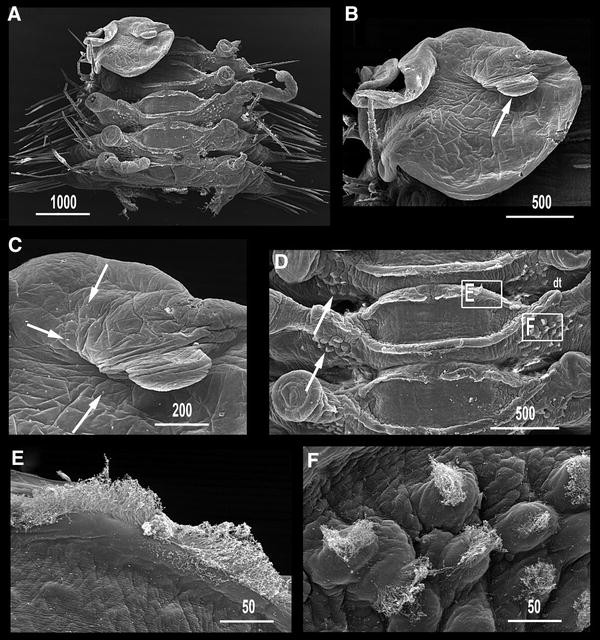
Medioantenna variopinta sp. nov. SEM micrographs. a. Mid-body segments, dorsal view. b. Whole view of elytron; arrow points to finger-like macropapilla. c. Detail of elytral surface around finger-like macropapilla; arrows point on micropapillae. d. Close view of body dorsum; arrows point to ciliated parapodial papillae; dt: dorsal tubercles from mid-body cirrophorous segment; squares mark the position of (e, f). e. Detail of dorsal ciliated crests. f. Detail of ciliated parapodial papillae. Scale bars are in μm
Fig. 7 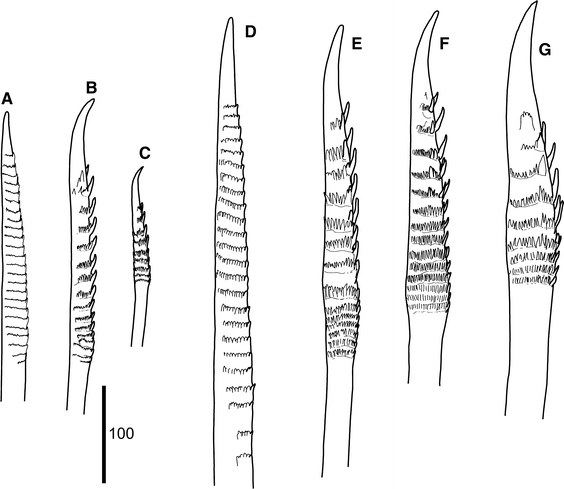
Medioantenna variopinta sp. nov. Holotype. a. Notochaeta from chaetiger 2. b. Dorsal-most neurochaeta from chaetiger 2. c. Ventral-most neurochaeta from chaetiger 2. d. Notochaeta from chaetiger 39. e. Dorsal-most neurochaeta from chaetiger 39. f. Mid-dorsal neurochaeta from chaetiger 39. g. Ventral-most neurochaeta from chaetiger 39. Scale bar is in μm
Fig. 8 
Medioantenna variopinta sp. nov. SEM micrographs. a. Mid-dorsal neurochaeta from mid-body. b. Detail of serration from the same mid-dorsal neurochaeta. c Detail of serration of a ventral neurochaeta from mid-body. d. Neurochaeta from mid-body. e. Detail of the serration of same neurochaeta. Scale bars are in μm
Fig. 9 
Medioantenna variopinta sp. nov. a–c. Underwater photographs of M. variopinta sp. nov. on unfertile S. secunda colonies (Photo Massimo Boyer); the polychaete is more evident when hydroid lacks the orange gonophores (a–b) or the hydranths (c). d, e. Stereomicroscope pictures of fertile specimens of S. secunda hosting M. variopinta, the worm livery perfectly mimics the host colors. White arrows: hydranths; black arrows: gonophores. Scale bars are in cm
-
Medioantenna clavata—Nishi and Tachikawa 1999: 107–110, Fig. 1b–j








Examined material
Holotype: female, cs in two fragments MNCN 16.01/13202, on a colony of Solanderia secunda, Celebes Sea, North Sulawesi, Mapia, 1°37′36.28″N; 124°48′8.46″E, 25 m deep, July 7, 2008. Paratype 1: female, cs with pharynx cut off for examination MNCN 16.01/13203, on the same host colony as the holotype, Celebes Sea, North Sulawesi, Mapia, 1°37′36.28″N; 124°48′8.46″E, 25 m deep, July 7, 2008. Paratype 2: juvenile, cs MNCN 16.01/13204, on S. secunda; Celebes Sea, North Sulawesi, Siladen Onong, 1°27′31.16″N; 124°45′59.53″E, 23.2 m deep, July 27, 2008. Additional material: mf with regenerated pygidial segments MNCN 16.01/13205, on S. secunda; Celebes Sea, North Sulawesi, Siladen Onong, 1°27′31.16″N; 124°45′59.53″E, 23.2 m deep, July 27, 2008.
Diagnosis
Long worm with 78 chaetigerous segments, and 34–38 pairs of elytra, markedly irregular in outline, with large fold on external margin, large finger-like papilla on posterior edge, and a few micropapillae irregularly distributed on dorsal surface. Neurochaetae all unidentate. Body orange, with prominent dark violet pigmentation on prostomium, and a combination of dark violet (at basis) and whitish (at tips) pigmentation in cephalic and body appendages and parapodial lobes, visible also in preserved specimens.
Description (based on holotype)
Prostomium bilobed, wider than longer, with small cephalic peaks. Two pairs of large dark brown eyes, ovate or round in shape; anterior pair larger than posterior one, lateral in position, on widest part of prostomium; posterior pair smaller than anterior one, near posterior margin of prostomium, dorsally orientated (Figs. 2a, b, 3a). Median antenna with large ceratophore, inserted antero-dorsally on prostomium; style long, smooth, cylindrical, distally slightly inflated, ending in short terminal filiform tip, with an internal axis (visible by transparency). Lateral antennae including ceratophores about six times shorter than median antenna, inserted termino-ventrally on distal prostomial end; styles about eight times shorter, basally inflated, smooth, with filiform tips. Palps smooth, stout, conical, gradually tapering to filiform tips, slightly shorter than median antenna (Figs. 2a, b, 3a, b). Facial tubercle prominent; mouth surrounded by two lateral lips, one dorsal with five lobes, the ventral lip large, lobular, with 10–12 poorly distinct lobes (Fig. 3b). Pharynx not everted, with four light brown jaws, all similar in shape and size (Paratype MNCN 16.01/13203). First segment not visible dorsally; tentaculophores moderately long, achaetous, with two pairs of tentacular cirri, similar in shape to median antenna, dorsal cirrus 1.5 longer than median antenna and twice as long as ventral cirrus (Figs. 2a, b, 3a), both with a few small papillae. Second segment with a semi-lunar nuchal lobe, carrying first pair of elytra dorsally (Fig. 2a, b); parapodia biramous; ventral cirri with style twice longer as in following segments, similar in shape to antennae and dorsal tentacular ones, with a few minute papillae (Fig. 4a); second and third ventral cirri similar in shape to first one, slightly decreasing in length, with only traces of violet pigmentation; successive ones abruptly becoming more than three times shorter, neither distally swollen nor pigmented, with papillae more numerous than on first one (Figs. 3h, i, 5a, b).
Elytra numbering 35 (right side) and 34 (left side), on chaetigers 2, 4, 5, 7 and alternating through chaetigers 23, 26, 29, 32, 35, 36, 38, 39, 42; thereafter asymmetrically arranged on 43, 46, 50, 53, 54, 57, 60, 63, 65, 67, 68, 69, 71, 74, 77 (right side) and 46, 47, 50, 53, 54, 57, 60, 61, 63, 65, 69, 71, 74, 76 (left side); reaching posterior end, overlapping antero-posteriorly, leaving mid-dorsum uncovered (Figs. 2a, b, 3c). All elytra basically oval, markedly irregular in outline, with a large fold on external margin, a very large finger-like papilla on posterior edge, and delicate transparent micropapillae scattered on dorsal surface; elytra basically transparent, with a few, small white spots, more abundant near edges; elytral nerves visible as ramifying whitish structures (Figs. 3f, g, 6a–c). Cirrigerous segments with dorsal tubercles (Fig. 6a, d). Dorsal cirri on postero-dorsal side of parapodia with cylindrical, relatively long cirrophores; styles smooth, cylindrical, with inflated subterminal ends, short filiform tips and few scattered small papillae, more distinct basally than distally (Fig. 3d, e). All segments with two prominent, transverse, dorsal ciliary bands, joining at basis of elytrophores or dorsal tubercles (Figs. 2a, 6a, d, e). Basal part of parapodia with numerous, prominent papillae bearing tufts of cilia on tips (Figs. 3h, i, 5c, 6d, f).
Parapodia relatively long, biramous (Figs. 3h, 4a–c); notopodia small, conical. Neuropodia with long anterodorsal acicular lobe (Fig. 5e); neuropodia deeply incised dorsally and ventrally, with long sub-triangular prechaetal lobe and digitiform supra-acicular process; postchaetal lobe half as long as prechaetal one, distally rounded; tips of noto- and neuroacicula protruding (Fig. 4a–c). Notochaetae as stout as or stouter as neurochaeta, arranged in a tuft (Figs. 2b, 3h), 6–8 per bundle, straight, with entire tips and slight serrations (Figs. 5c, 7a, d, 8d, e). All neurochaetae of same type, falcate, with distinct rows of spines, with distally bent, unidentate tips; 18–20 per bundle (Figs. 5d, 7b, c, e–g, 8a–c).
Nephridial papillae from segment 6 to posterior end, becoming elongated, upwardly directed, nearly reaching upper side of notopodia in segments 16–18 (Fig. 5g). Pygidium with terminal anus; two ventral cirri below anus, relatively long, as long as and similar in shape to posterior-most dorsal cirri.
Measurements and variability
Holotype (cs), MNCN 16.01/13202. L: 44 mm, WW: 1.5 mm, WP: 4.1 mm, 78 segments. Paratypes: MNCN 16.01/13203 (cs), L: 34 mm, WW: 1.6 mm, WP: 3.3 mm, 77 segments; MNCN 16.01/13204 (cs, juvenile), L :11 mm, WW: 0.7 mm, WP: 1.9 mm, 43 segments. Additional material: MNCN 16.01/13205 (mf, with regenerated posterior segments), L: 6 mm, WW: 1.8 mm, WP: 4.0 mm, 8 segments.
Number of elytra varying both between left and right sides and between worms. The juvenile paratype MNCN 16.01/13204 has 18 regularly distributed pairs (Table 2). The adult elytral distribution is regular up to segments 42 (Holotype) and 35 (Paratype MNCN 16.01/13203), and the total number of elytra on left and right sides also differed: 35/34 (holotype) and 36/29 (Paratype MNCN 16.01/13203) (Table 2). The number of notochaetae per bundle do not vary significantly in animals of different size (6–8 in holotype and juvenile), while the neurochaetae are less numerous in large specimens (holotype 11–13, juvenile 18–20). The juvenile has both eye pairs nearly equal in size, instead of the anterior pair larger than posterior one typical of adults. In addition, the holotype and the mid-body fragment have elongated nephridial papillae, which were much shorter in the other paratypes.
Coloration
Body bright orange on living worms, with a bright violet pigmentation on prostomium, parapodia, cirrophores and basal parts of antennae, palps, dorsal cirri and first 2–3 pairs of ventral cirri. Distal ends on cephalic appendages and dorsal cirri bright whitish. The orange body mimics the large orange eumedusoids of the host colony, while the white pigmentation on the cephalic appendages and on dorsal cirri and the finger-like macropapillae of the elytra mimic the coloring of the host polyps (Fig. 9a–e). Elytra with whitish spots near the margin, whitish pigmentation on distal part of the finger-like papilla, and a pale-white ring surrounding the area of attachment to the elytrophore, extending to the nearby surface (Fig. 3f, g). Mimicry of worms on fertile hydroid colonies (Fig. 9d, e) is more efficient than on unfertile colonies (Fig. 9a, b).
Body of alcohol-preserved worms becoming whitish in background, with prominent dark violet pigmentation on prostomium, parapodia, cirrophores and basal parts of antennae, palps, dorsal cirri and first 2–3 pairs of ventral cirri (Fig. 3a–d).
Etymology
The specific epithet variopinta refers to one of the most distinctive features of the species, which is its spectacular bright orange, whitish and violet color pattern that perfectly mimics the fertile colonies of its host hydroid.
Taxonomic remarks
The morphology of the new species fits well with the diagnosis of Medioantenna, as proposed by Imajima (1997) for M. clavata. Both species have an enlarged ceratophore on the median antenna, arising antero-dorsally from the prostomium, and lateral antennae with small ceratophores. The elytral distribution coincided from segments 2 to 35 and became irregular on posterior-most segments (Table 2). However, M. variopinta sp. nov. has a very large finger-like papilla on the rear region of elytra, clearly separated from the margin instead of a soft lump on the rear margin of the elytra in M. clavata (Imajima 1997, Fig. 7e, d). Furthermore, the strongly serrated neurochaetae are similar in the two species, but all of them have unidentate tips in M. variopinta sp. nov., instead of bidentate tips in segment two in M. clavata (Imajima 1997; Fig. 9g, h). The notochaetae are 6–8 per bundle in the new species and are more numerous in M. clavata. Although the exact number was not reported in the original description, they ranged from 11 to 16 per bundle according to the illustrations (Imajima 1997; Figs. 7d, 8a, b) and from 13 to 16 per bundle in observed paratype. Additionally, M. variopinta sp. nov. is slender and has more segments than M. clavata. The holotypes of both species have nearly the same length (i.e., 44 and 45 mm, respectively), but M. variopinta sp. nov. is substantially more slender: 4.1 mm WP versus ca. 6.0 mm WP in M. clavata (based on paratype measurements). In addition, the chaetigers are more numerous in the former (up to 77–83) than in the latter (up to 59) (Table 2). Additional minor differences are the presence of micropapillae on the elytral surface in M. variopinta sp. nov., which are absent in M. clavata, and the prominent dark violet pigmentation on the prostomium and basal parts of prostomial and body appendages visible in M. variopinta sp. nov. even in preserved specimens, which is absent in M. clavata.
The specimens from Ogasawara Islands, described by Nishi and Tachikawa (1999) as belonging to M. clavata, deserve special attention. Their morphology and coloring agree with M. variopinta sp. nov. but there seem to be several differences. The tentacular segment bears chaetae, the prostomium lacks cephalic peaks, the second segment lacks nuchal fold, the anal cirri are absent, the elytra lack micropapillae, and the dorsal ciliary bands are absent.
Among them, the first one is the most intriguing feature, as chaetae are absent from the tentacular segment in both in Imajima (1997) and in our Indonesian worms, and also because having achaetous tentaculophores is one of the diagnostic features of the Arctonoinae (Hanley 1989; Wehe 2006). Cephalic peaks are not prominent in our specimens and were described as “rudimentary” in the Medioantenna generic diagnosis (Imajima 1997). As the shape of cephalic peaks might be affected by fixation and may even vary among species of the same genus (e.g., in Hololepidella) or among specimens of the same species (e.g., in Hololepidella laingensis, Britayev et al. 1999), we consider that its absence may not have a great significance for the identification of species within this genus. In turn, the absence of the nuchal fold and anal cirri in the specimens described by Nishi and Tachikawa (1999) could be a consequence of either an intra-specific variation or a fixation bias (nuchal fold) and of a loss during sample handling (anal cirri). Finally, the presence of the two dorsal ciliary bands, clearly visible in our specimens, has been reported neither by Imajima (1997) nor by Nishi and Tachikawa (1999). Despite this, the position of the inter-segmentary lines on Fig. 1a, b in Nishi and Tachikawa (1999) perfectly agrees with that of the ciliary bands, therefore being present (but not reported) in the worms from Ogasawara Islands.
Since the above-mentioned differences might be caused by sample handling or observation biases, they are not sufficient to consider both the Japanese and Indonesian specimens as different species. Keeping this in mind, and taking into account that we did not have the opportunity to check the specimens from Ogasawara Islands, both populations appear to coincide in virtually all diagnostic characteristics (including appendage and elytral morphology, color pattern, mode of life and likely host species), leading us to consider that both belong to Medioantenna variopinta sp. nov.
Biology
Medioantenna variopinta sp. nov. was found living on colonies of Solanderia secunda at 12–25 m deep (Fig. 9a–e). The symbionts were not present on every checked colony, but their occurrence could have been underestimated due to their ability to quickly hide themselves among the intricate basal branches when the colonies are disturbed. The number of infested hydroid colonies rarely exceeded 3–5%, but exceptionally reached 50% (Table 1). The intensity of infestation varied from 1 to 3 worms per colony. Four of five infested colonies harbored 2 or 3 worms each (Table 1), possibly indicating pair formation.
Damaged and regenerating elytra and dorsal cirri were found on all specimens (for instance: holotype—2, 4, 7, 9, 11, 13, 15 chaetigers right side, 4, 5, 7, 9, 13 chaetigers left side; Paratype MNCN 16.01/13203–7 chaetiger left side, Paratype MNCN 16.01/13204–32 both sides, 35, 38, 39—right side, Fig. 6d), which may indicate the possibility of intra-specific aggressive interactions. In the MNCN 16.01/13205 specimen (fragment), the few posterior-most segments, pygidium and anal cirri were clearly regenerating, which may indicate the presence of predator attacks, as demonstrated in other scale-worms (Britayev 1991; Britayev and Zamyshliak 1996).
The holotype and the fragment with regenerating posterior end were females and had elongated nephridial papillae, while paratype 1 (found on the same hydroid colony as holotype) seemed to be a male and had short nephridial papillae. The single juvenile found also had short papillae.
Distribution
Siladen (Type Locality), Bunaken and Mapia from North Sulawesi, Indonesia; Ogasawara Islands, Japan.
-
Phylum CNIDARIA
-
Class HYDROZOA
-
Family SOLANDERIIDAE Marshall 1892
-
Genus Solanderia Duchassaing and Michelin, 1846 = Chitina Carter, 1873
-
Solanderia secunda (Inaba 1892 ) (Figs. 5, 10)
Fig. 10 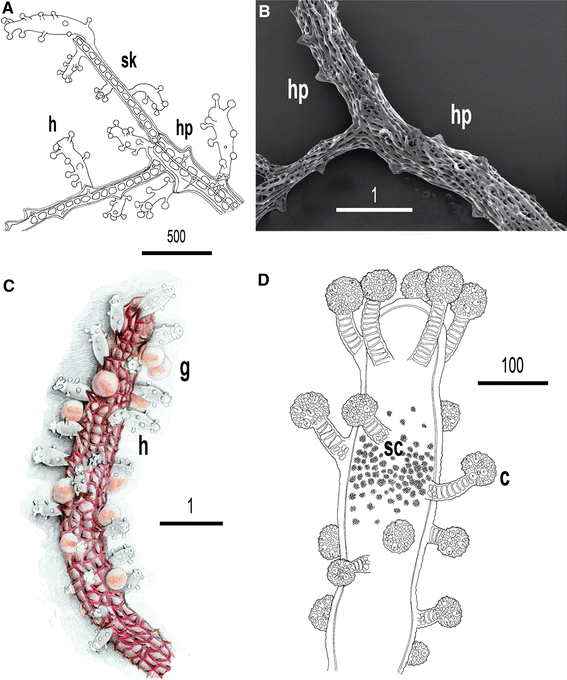
Solanderia secunda. a. Drawing of a colony portion showing hydranths (h) and hydrophores (hp). The inner skeleton is fenestrated (sk). b. SEM picture of the skeleton of S. secunda. Hydrophores (hp) and skeletal meshes are clearly visible. c. Portion of a branch showing hydranths (h) and gonophores (g). d. Particular of a hydranth with capitate tentacles and cnidocysts inside capitula (c); at middle of polyp sphaerolous and refringent cells are visible (sc). Scale bars of a and d are μm; scale bars of b, c are in mm
-
Dendrocoryne secunda Inaba 1892:98, figs. 111–113
-
Solanderia secunda—Bouillon,Wouters and Boero 1992: 12, pls 5–6, 10–12, synonymy

Material examined
Celebes Sea, North Sulawesi, Indonesia: about thirty colonies on the fringing reef of Bunaken Timur, 1°36′34.20″N; 124°46’57.96″E, 10–30 m deep, August 1, 2007 and October 10, 2008; two colonies on the patch reef of Mapia, 1°37’36.28″N; 124°48’8.46″E, 25 m deep, October 7, 2008; two colonies on the fringing reef of Negeri, 1°36’57.11″N; 124°42’15.63″E, 10 m deep, July 24, 2008; about twenty colonies on the fringing reef of Siladen Jeti, 1°37’39.10″N; 124°48’0.26″E, 10–30 m deep, February 11, 2008 and November 27, 2008; about twenty colonies on the fringing reef of Siladen Onong, 1°27’31.16″N; 124°45’59.53″E, 10–30 m deep, July 27, 2008 and February 26, 2005; one colony on the fringing reef of Tanjung Pisok, 1°33’59.00″N; 124°47’55.61″E, November 20, 2006.
Diagnosis
Very large colonies, up to 30 cm high and 50–500 cm2 in surface, erect, fan-shaped, with one to several fascicled stems; branch thickness decreasing toward distal extremities (Figs. 9a,10a). Ramifications always on a plane. Skeleton permeated, covered by living tissue (coenosarc). Surface of basal part of stems almost smooth, lacking hydrophores (triangular skeletal structures at basis of hydranths). Central and peripheral portions of colonies with thinner branches bearing prominent hydrophores; branches rounded or flattened, ribbon-like.
Hydranths pin-shaped, flanked by hydrophores, with an oral whorl of 4–5 capitate tentacles and 13–15 scattered aboral capitate tentacles. Sphaerolous and refringent cells grouped in a dense cluster at middle of polyp (Fig. 10d). Nematocysts as stenoteles with two sizes (11,5–14 × 15–18 μm and 5–7 × 7–9 μm); inside capitula. Living polyps white, able to curve after mechanical stimulation.
Gonophores are eumedusoids with four radial canals, a circular canal and spadix; spherical or oval, supported by a short pedicel; gonophores mostly concentrated in central part of colonies, on secondary branches.
Coloration
Exoskeleton varying from dark red (on thicker parts) to brilliant red (on thinner branches); hydranths white and gonophores bright orange; canals and spadix reddish (Figs. 9a–d, 10c).
Biology
Solanderia secunda were found on coral reefs at Bunaken Marine Park, as well as on scattered corals in the sandy slope at Mapia House Reef. The colonies were most abundant along the vertical coral walls. The highest colony density was found at Siladen Jeti, Siladen Onong and Bunaken Timur, where the colonies are under the constant influence of strong, bi-directional tidal currents. Colonies were rare in sites with high sedimentation rates, such as Tanjung Pisok where high turbidity discourages solanderiid development despite the strong currents. Colonies were generally oriented perpendicularly to the reef wall, at 10–35 m deep, being most common at a depth between 10 and 20 m. The species is fertile all over the year, except July.
Distribution
Widely distributed in the tropical Indo-West Pacific (Bouillon et al. 1992; Di Camillo et al. 2008; Kirkendale and Calder 2003; Millard 1975; Schuchert 2003; Vervoort 1966; Watson 1999).
Characteristics of the association between Medioantenna variopinta sp. nov. and Solanderia secunda
Despite the presence of two Solanderia species in the study area, S. procumbens and S. secunda, Medioantenna variopinta sp. nov. was only observed in association with the latter. The polychaetes were found on colonies living at a depth between 12 and 25 m, either on the large fan-shaped portion or hidden in the basal portion, where branches formed an irregular bundle (Fig. 9a–e). The number of polychaete specimens present on the same colony ranged from one to a maximum of three (Table 1).
The color pattern of the symbiotic scale-worm closely mimicked that of the fertile specimens of S. secunda. Dorsal cirri resembled the hydranths both in shape and in color. The basal part of each cirri had a V-shaped, dark violet pigmentation resembling that of hydrophores, while the apical portion resembled the hydranths, with its pin-shape and numerous white spots (Fig. 9d, e). Even though cirri lack structure similar to the hydroid tentacles, the scattered white spots mimicked their irregular distribution, a camouflage mimicry that is reinforced by the white spots scattered on the transparent elytrae. The white finger-like macropapillae on elytra resemble the hydranth’s hypostome, the dark violet color of parapodia matched that of hydroid’s skeleton, and the bright orange body resembled the gonophores (Fig. 9d, e). The camouflage turns out to be most effective on fertile colonies bearing the large orange eumedusoids, while the worm’s orange body seems to be more distinguishable from the background when the hydroid is unfertile (Fig. 9a–c). Although S. secunda is fertile almost all year round (Di Camillo et al. 2008), during the very short periods that the hydroid lacks gonophores, M. variopinta sp. nov. stands out on the host, probably becoming more vulnerable to predator attacks (Fig. 9a, b). However, it must be taken into account that we described (and figured) the worm and host coloring under artificial conditions (i.e., in the laboratory or under bright lighting). In the native underwater conditions, the apparently evident differences in color between the symbiont and the host might not be perceived and the pattern resulting from the contrasts in pigmentation intensity (i.e., cirri and elytra vs. body in the symbiont, hydrants and gonophores versus exoskeleton in the host) becomes even more relevant than the colors themselves. This gives rise to an effective camouflage and the two partners appear as a unique organism, particularly in ripe hydroid colonies (even under artificial lighting).
In natural conditions, scale-worms crawl slowly among the hydrozoan branches. When disturbed, they quickly run down to the basal part of the colony, sheltering among the sessile epibionts that can be found in this region. When detached from the host, the scale-worms may autotomise. When this happens, their anterior ends quickly swim toward the host. Chemical cues released by hosts to attract the symbionts (i.e., the so-called host factor) have been previously reported or suggested for highly specific associations (see Martin and Britayev 1998 and references herein). In the case of M. variopinta sp. nov., the behavior of the autotomised fragments also suggests the presence of such a host factor. Moreover, entire worms were also always able to find the fragments of hydroid colonies placed in the same container, particularly if the host was the same individual that had harbored the specimen. However, we did not perform real-choice experiments. Thus, although we strongly suspect the presence of a chemical cue causing the response, we could not distinguish whether the observed behavior corresponded to a host-factor response or simply to a random encountering or sheltering.
On the other hand, Medioantenna variopinta sp. nov. has never been seen feeding on polyps or gonophores of its host. So we suspect that scale-worms may prey on other vagile animals living on the same colonies, such as amphipods, copepods or ophiuroids. It is interesting to note that, on the colonies, scale-worms are often seen rubbing the chaetae of adjacent parapodia. However, we cannot assess whether this results from a self-cleaning behavior (either triggered by any host-released substance or triggered by a natural accumulation of debris on chaetae) or from a host-cleaning behavior.
Discussion
From a morphological point of view, Medioantenna variopinta sp. nov. is well characterized by its elytra which have a large finger-like papilla on their posterior edge, their unidentate neurochaetae and their peculiar color pattern. However, a couple of features of the species seem to be quite rare among scale-worms: the extremely high variability in elytral distribution (along the body in different specimens and between the sides in a given specimen) and the elongated, upwardly directed nephridial papillae (found only in females). The variability in elytral distribution characterizes both M. clavata and the Japanese and Indonesian specimens of M. variopinta sp. nov. (Table 2). This variability is rare among scale-worms and has only been reported for other long-bodied species within Arctonoe or Gastrolepidia, where the irregularity and asymmetry of elytra starts on chaetiger 33–36 or later (Hanley 1989; Uschakov 1982), or within Enipo and Neohololepidella (Muir 1982). Gorekia, Scalisetosus and Leucia are not so long and thin, but all have non-standard elytral arrangements (Muir 1982). In M. variopinta sp. nov., the distribution was regular up to chaetiger 35 (our specimens) or 22 (Japanese specimens) (Table 2).
As for the nephridial papillae, they are usually relatively small and ventro-laterally directed among scale-worms (Uschakov 1982). Elongated and dorsally directed papillae have only been reported once, in ripe Harmothoe imbricata females, where this peculiar position between parapodia was adopted to deliver eggs in a dorsal position, to be then brooded beneath elytra (Daly 1972). After breeding, the papillae regressed to a near-juvenile (or male) condition (Daly 1972). Therefore, our finding suggests a pre-reproductive or reproductive status for the females of M. variopinta sp. nov. Indirectly, this also supports the existence of a regular distribution with pair formation which was also reported for H. imbricata (Daly 1972). Eventually, our findings suggest the possibility of brooding in M. variopinta sp. nov., although it has not been possible to confirm this hypothesis.
The host Solanderia secunda, in turn, is the largest anthomedusan hydroid in Bunaken among those forming a thick, anastomosed perisarcal skeleton. However, the colonies do not harbor many organisms. The vagile (i.e., copepods, amphipods, ophiuroids and gastropods)- and sessile (i.e., pteroid bivalves on the fan, barnacles and bryozoans at the basal region)-associated faunas are very poor, the latter likely because larvae are not able to settle on the living coenosarc covering the skeleton. In addition, several natural products (i.e., solandelactones) have been isolated from S. secunda that have an antifouling activity preventing larval settlement (Seo et al. 1996). Medioantenna variopinta sp. nov. is probably immune to these natural products so its larvae can settle, and adults can spent their whole life, on the hydroid surface. In addition to help in removing debris from chaetae, the observed rubbing behavior could also be induced by an accumulation of mucus from the host. This accumulation occurring in the locomotive appendages (i.e., the parapods) could certainly limit the moving ability of the symbiont, which may feel the necessity to eliminate the excess of mucus. However, it is also possible that the worm could take advantage of a thin layer of mucus being deposited on its body. This layer may contain some of the hydroid’s natural products that may play a deterrent role helping the worm to avoid predation. Although this is an interesting behavioral consequence of the association, further studies are required to obtain additional evidence. In addition to this possible benefit for the symbiont, the regular presence of M. variopinta sp. nov. on the colonies suggests by itself an intimate relationship with the host hydroid which is additionally enhanced by the cryptic coloring and mimicry of the scale-worm.
Nishi and Tachikawa (1999) found the Ogasawara specimens on a solanderiid, reported as Solanderia misakiensis in the abstract and then referred to as S. secunda in the text. The most relevant difference between the two species is the absence of hydrophores in the former, but Nishi and Tachikawa (1999) did not supply a description for the hydroid so that its classification is, at least, doubtful. In case, the hydroid from Ogasawara was indeed S. misakinensis that would mean Medioantenna variopinta sp. nov. is able to live on two similar solanderiid species. Both hydroids have dark colonies and probably produce similar chemicals, which could not discourage the presence of the symbiont. However, it is more likely that the Japanese hydroid also belongs to S. secunda. In fact, this species has often been reported from Japanese waters (Bouillon et al. 1992), and even its original description by Inaba (1892) has Misaki (Japan) as type locality. Thus, it seems more feasible to consider the association as species-specific. In turn, this specificity is now confirmed by the fact that, in Bunaken, M. variopinta sp. nov. were never observed on the yellow colonies of S. procumbens, which lived exactly in the same area and habitat as the infested colonies of S. secunda. Nevertheless, we have not been able to precise the real nature of the relationship between the new species of scale-worm and its hydroid host.
More than 300 polychaete species are known to be involved in different types of symbiotic relationships with other marine invertebrates which consequently developing morphological, behavioral, physiological or reproductive adaptations to their particular modes of life (Martin and Britayev 1998; Martin et al. 2002). In monoxenous species, it often happens that the shape of some symbiont appendages perfectly mimics a relevant morphological characteristic of the host. For instance, the swollen dorsal cirri of the scale-worm Asterophylia carlae mimic the ambulacral feet of its host starfishes (Hanley 1989; Martin and Britayev 1998). Also, the whole body may display camouflage colorings exactly matching the host color such as those of the syllids Haplosyllis chamaeleon and H. villogorgicola and their respective gorgonian hosts (Laubier 1960, 1968; Martin et al. 2002), or Alcyonosyllis phili and its alcyonacean host (Glasby and Watson 2001). In particular, it must be pointed out that A. phili has a color pattern which closely resembles that of M. variopinta sp. nov. In turn, polyxenous species may change their color in relation to that of the hosts, such as in the case of the syllid Branchiosyllis oculata, associated with sponges, or the scale-worm Gastrolepidia clavigera, associated with holothuroids (Pawlik 1983; Britayev and Lyskin 2002). Some sponge-associated syllids feed on host tissues showing different colors that mimic their particular host (Pawlik 1983; Magnino and Gaino 1998). Among gorgonian-associated syllids, in turn, either feeding on the same food as the host, kleptoparasitism (i.e., stealing food from the gastral cavities of the host polyps) and/or co-evolution seems to be the reason for that mimicry (Martin et al. 2002). However, syllids and scale-worms differ in their feeding behavior, the former suck food by means of their pharynx and their muscular proventricle, while the latter bite their prey with the help of their four powerful parrot’s beak-like jaws. Curiously enough, among the few cases when feeding of symbiotic scale-worms was studied (e.g., Arctonoe vittata, Gastrolepidia clavigera), a wide spectrum of strategies, from host-cleaning, stealing food, foraging on free-living organisms to feeding on host tissues, were demonstrated (Wagner et al. 1979; Britayev et al. 1989; Britayev and Lyskin 2002).
The study of feeding strategies also revealed complementary behavioral trends. For instance, in addition to their prey, both A. vitatta and G. clavigera had chaetae of conspecifics inside the gut whose presence was associated with that of traumas (i.e., on parapodia, head, elytra and body appendages) exactly as those found in M. variopinta sp. nov. These findings have been considered as a demonstration of aggressive territorial behavior in symbiotic (Britayev 1991; Britayev and Lyskin 2002) as well as in free-living (e.g., Plyuscheva et al. 2010) scale-worms, often leading to regular distributions in the former (Martin and Britayev 1998), and our new species seems to agree in both distribution pattern and behavior.
The few well-known host–symbiont relationships appear to be more complex than it was initially thought, with the associations often resulting to be mutualistic rather than simple parasitic or commensalistic. In most cases, the misinterpretation was caused by both an initial lack of knowledge and our usual antropogenic way of interpreting what we are looking at. For instance, the histriobdellid Histriobdella homari was reported as an ectoparasite of the branchial chambers of crustaceans (e.g., in Paris 1955 or Clark 1956), mostly because of its leech-like shape, and was later confirmed as a host-cleaning mutualist (Jennings and Gelder 1976). Also, Eunice norvergica, a gall-forming eunicid (an a priori parasitic behavior), enhanced the growth of their cold-water corals hosts by favoring the joining of colonies and the development of reef patches in deep waters (Roberts 2005). Among scale-worms, Arctonoe vitatta was considered to be a commensal of their limpet or starfish hosts and later re-considered as a mutualist, which is able to defend the former from the attacks of predatory starfishes (Dimock and Dimock 1969; Williams 2009) or to keep the latter clean by replacing the normal function of the pedicellaria, lacking in their starfish hosts (Wagner et al. 1979). This last partnership was also confirmed as mutualism from a chemical point of view, as both the symbiont and the host were attracted by cues coming from the host and the symbiont, respectively (Wagner et al. 1979).
Despite the numerous examples of symbiotic polychaetes reported to date, their behavioral and reproductive strategies, as well as the origin of their food and the nature of their cryptic coloring, are still open questions in most relationships involving polychaetes, including scale-worms (Martin and Britayev 1998). The main reasons are probably the rarity of the most known associations, but also the difficulties of either observing the partnership in native conditions or keeping them in experimental tanks for long periods, as it happens with our new species. Therefore, all observed details on any given association may be relevant and must be clearly reported, even if it initially leads to wrong conclusions, such as the report of the Ogasawara Islands specimens as Medioantenna clavata by Nishi and Tachikawa (1999). Further studies will certainly help to understand the real nature of either the associates or their associations, as well as to assess the complex relationships between symbiotic polychaetes and their hosts, and again Medioantenna variopinta sp. nov. is not an exception.
References
Barnich R, Fiege D (2001) The Mediterranean species of Malmgreniella Hartman, 1967 (Polychaeta: Polybnoinae), including the description of a new species. J Nat Hist 35:1119–1142
Barnich R, Fiege D (2003) The Aproditoidea (Annelida: Polychaeta) of the Mediterranean Sea. Abh Senckenberg Naturforsch Ges 559:1–167
Boero F, Bouillon J (2005) Cnidaria and Ctenophora. In: Rhode K (ed) Marine parasitology. p, CSIRO Collingwood, pp 177–182
Bouillon J (1994) Embranchement des Cnidaires (Cnidaria). In: Grassé PP, Doumec D (eds) Traité de Zoologie, vol 3, pp 1–28
Bouillon J, Boero F (1987) The life cycle of Teissiera medusifera (Teissieridae, Anthomedusae, Hydrozoa, Cnidaria). Indo-Malayan Zool 1:1–9
Bouillon J, Wouters K, Boero F (1992) Étude des Solanderiidae de la Baie de Hansa (Papouasie Nouvelle-Guinée) avec une révision du genre Solanderia (Cnidaria, Hydrozoa). Bull Inst r Sci nat Belg Biol 62:5–33
Britayev TA (1981) Two new species of commensal polynoids (Polychaeta: Polynoidae) and bibliography on polychaetes, symbionts of Coelenterata. [In Russian]. Zool Zh 60:817–824
Britayev TA (1991) Life cycle of the symbiotic scale-worm Arctonoe vittata (Polychaeta: Polynoidae). In: Systematics, biology and Morphology of World Polychaeta. Proceedings of the 2nd International Polychaeta Conference. Ophelia Suppl 5:305–312
Britayev TA, Lyskin SA (2002) Feeding of the symbiotic polychaete Gastrolepidia clavigera (Polynoidae) and its interactions with its hosts. Doklady Biol Sci 385:352–356
Britayev TA, San Martín G (2001) Description and life-history traits of a new species of Proceraea with larvae infecting Abietinaria turgida (Polychaeta, Syllidae and Hydrozoa, Sertulariidae). Ophelia 54:105–113
Britayev TA, Zamyshliak EA (1996) Association of the commensal scaleworm Gastrolepidia clavigera (Polychaeta: Polynoidae) with holothurians near the coast of South Vietnam. Ophelia 45:175–190
Britayev TA, Smurov AV, Adrianov AV, Bazhin AG, Rhzavsky AV (1989) Ecology of symbiotic Polychaete Arctonoe vittata according to the peculiarity of starfish Asterias ratbunae ecology. In: Sveshnikov VA (ed) Symbiosis among marine animals. A.N. Severtzov Institute, Russian Academy of Sciences Moscow, Moscow, pp 102–127
Britayev TA, Doignon G, Eeckahaut I (1999) Symbiotic polychaetes from Papua New Guinea associated with echinoderms, with descriptions of three new species. Cah Biol Mar 40:359–374
Cairns SD (1987) Conopora adeta, a new species (Hydrozoa: Stylasteridae) from Australia, the first known unattached stylasterid. Proc Biol Soc Wash 100:141–146
Clark RB (1956) Capitella capitata as a commensal, with a bibliography of parasitism and commensalism in the polychaetes. Ann Mag Natur Hist 9:433–448
Daly JM (1972) The maturation and breeding biology of Harmothoe imbricata (Polychaeta: Polynoidae). Mar Biol 12:53–66
Di Camillo CG, Bavestrello G, Valisano L, Puce S (2008) Spatial and temporal distribution in a tropical hydroid assemblage. J Mar Biol Ass UK 88:1589–1599
Dimock RV Jr, Dimock JG (1969) A possible “defense” response in a commensal polychaete. Veliger 12:65–68
Galea HR, Försterra G, Häussermann V (2007) Additions to the hydroids (Cnidaria: Hydrozoa) from the fjords region of southern Chile. Check List 1650:55–68
Genzano GN, San Martín G (2002) Association between the polychaete Procerastea halleziana (Polychaeta: Syllidae: Autolytinae) and the hydroid Tubularia crocea (Cnidaria: Hydrozoa) from the Mar del Plata intertidal zone, Argentina. Cah Biol Mar 43:165–170
Gidholm L (1966) A revision of Autolytinae (Syllidae, Polychaeta) with special reference to Scandinavian species, and with notes on external and internal morphology, reproduction and ecology. Ark Zool 19:157–213
Gili JM, Hughes RG (1995) The ecology of marine benthic hydroids. Oceanogr Mar Biol Annu Rev 33:351–426
Glasby CJ, Watson C (2001) A new genus and species of Syllidae (Annelida: Polychaeta) commensal with octocorals. Beagle Rec North territ Mus Art Sci 17:43–51
Hanley JR (1989) Revision of the scaleworm genera Arctonoe Chamberlin and Gastrolepidia Schmarda (Polychaeta, Polynoidae) with the erection of a new subfamily, Arctonoinae. Beagle Rec North territ Mus Art Sci 6:1–34
Hanley JR, Burke M (1991) Polychaeta Polynoidae: Scaleworms of the Chesterfield Islands and Fairway Reefs, Coral Sea. In: Crosnier A (ed) Résultats des Campaignes MUSORSTOM. Mém Mus natn Hist nat, (A), 151, Paris ISBN: 2-85653-186-5, pp 9–82
Hartmann-Schröder G (1992) Drei neue Polychaeten-arten der familien Polynoidae und Syllidae von Neu-Kaledonien, assoziiert mit einer verkalten Hydrozoe. Helgoländer Meeresunters 46:93–101
Hoeksema B (2007) Delineation of the Indo-Malayan centre of maximum marine biodiversity: The coral triangle. In: W Renema (ed) Biogeography, time, and place: Distributions, barriers, and islands, p 117–178 © 2007 Springer
Imajima M (1997) Polychaetous annelids from Sagami Bay and Sagami Sea collected by the Emperor Showa of Japan and deposited at the Showa Memorial Institute, National Science Museum, Tokio. Natn Sci Mus Monogr 13:1–138
Inaba M (1892) Soshu, Miura, Misaki ni oide edaru Hydroidea. [The hydroids collected at Miura and Misaki in Soshu.]. Zool Mag 4(93–101):124–131
Jennings JB, Gelder SR (1976) Observations on the feeding mechanism, diet and digestive physiology of Histriobdella homari van Beneden 1858: an aberrant polychaete symbiotic with North American and European lobsters. Biol Bull Mar Biol Lab Woods Hole 151:489–517
Kirkendale L, Calder D (2003) Hydroids (Cnidaria: Hydrozoa) from Guam and the commonwealth of the Northern Marianas Islands (CNMI). Micronesica 35–36:159–188
Lagardère F, Tardy J (1980) Un faciès d’épifaune nouveau: le faciès à Ectopleura dumortieri (van Beneden) et Electra pilosa (Linné) faune associée, cartographie et évolution saissoniére. Cah Biol Mar 21:265–278
Laubier L (1960) Une nouvelle sous-espèce de Syllidien: Haplosyllis depressa Augener sp. nov. chamaeleon, ectoparasite sur l’octocoralliaire Muricea chamaeleon Von Koch. Vie et Milieu 11:75–87
Laubier L (1968) Contribution à la faunistique du coralligène VII. À propos de quelques Annélides Polychètes rares ou nouvelles (Chrysopetallidae, Syllidae et Spionidae). Ann Inst Océanog Mónaco 46:79–107
Magnino G, Gaino E (1998) Haplosyllis spongicola (Grübe) (Polychaeta, Syllidae) associated with two species of sponges from east Africa (Tanzania, Indian Ocean). P.S.Z.N. I: Mar Ecol 19(2):77–87
Martin D, Britayev TA (1998) Symbiotic polychaetes: review of known species. Oceanogr Mar Biol Ann Rev 36:217–340
Martin D, Núñez J, Riera R, Gil J (2002) On the associations between Haplosyllis (Polychaeta, Syllidae) and gorgonians (Cnidaria, Octocorallaria), with the description of a new species. Biol J Linn Soc 77:455–477
Millard NAH (1975) Monograph on the Hydroida of Southern Africa. Ann S Afr Mus 68:1–513
Molodtsova T, Budaeva N (2007) Modifications of corallum morphology in black corals as an effect of associated fauna. Bull Mar Sci 81:469–480
Muir AI (1982) Generic characters in the Polynoinae (Annelida, Polychaeta) with notes on the higher classification of scale-worms (Aphroditacea). Bull Br Mus nat Hist 43:153–177
Nishi E, Tachikawa H (1999) New record of a commensal scale worm Medioantenna clavata Imajima, 1997 (Polychaeta: Polynoidae), from Ogasawara Islands, Japan. Nat Hist Res 5:107–110
Nygren A (2004) Revision of Autolytinae (Syllidae: Polychaeta). Zootaxa 680:1–314
Paris J (1955) Commensalisme et parasitisme chez les annèlids polychètes. Vie et Milieu 6:525–536
Pawlik JR (1983) A sponge-eating worm from Bermuda: Branchiosyllis oculata (Polychaeta, Syllidae). P.S.Z.N.I: Mar Ecol 4:65–79
Pettibone MH (1989) A new species of Benhamipolynoe (Polychaeta: Polynoidae: Lepidasteniinae) from Australia, associated with the unattached stylasterid coral Conopora adeta. Proc Biol Soc Wash 102:300–304
Pettibone MH (1991) Polynoid polychaetes commensal with antipatharian corals. Proc Biol Soc Wash 104:714–726
Plyuscheva M, Martin D, Britayev TA (2010) Diet analyses of the scale-worms Lepidonotus squamatus and Harmothoe imbricata (Polychaeta, Polynoidae) in the White Sea. Mar Biol Res 6(3):271–281
Puce S, Cerrano C, Di Camillo CG, Bavestrello G (2008) Hydroidomedusae (Cnidaria: Hydrozoa) symbiotic radiation. J Mar Biol Ass UK 88:1715–1721
Roberts JM (2005) Reef-aggregating behaviour by symbiotic eunicid polychaetes from cold-water corals: do worms assemble reefs? J Mar Biol Ass UK 85(4):813–819
Schuchert P (2003) Hydroids (Cnidaria, Hydrozoa) of the Danish expedition to the Kei Islands. Steenstrupia 27:137–256
Seo Y, Cho KW, Rho J-R, Shin J, Kwon B-M, Bok S-H, Song J-I (1996) Solandelactones A-I, lactonized cyclopropyl oxylipins isolated from the hydroid Solanderia secunda. Tetrahedon 52:10583–10596
Uschakov PV (1982) Polychaetes, Fauna of the USSR 2(1), Polychaetes of the Suborder Aphroditiformia of the Arctic Ocean and the Northwestern Part of the Pacific, Families Aphroditidae and Polynoidae, Academy of Sciences of the USSR, Zoological Institute vol. NAUKA, Moscow
Vervoort W (1966) Skeletal structure in the Solanderiidae and its bearing on hydroid classification. In: Rees WJ (ed) The Cnidaria and their Evolution. The Proceedings of a Symposium held at The Zoological Society of London on 3 and 4 March 1965, p 372–396
Wagner RH, Phillips DW, Standing JD, Hand C (1979) Commensalism or mutualism: attraction of a sea star towards a symbiotic polychaete. J Exp Mar Biol Ecol 39:205–210
Watson JE (1999) Hydroids (Hydrozoa: Anthoathecata) from the Beagle Gulf and Darwin Harbour, northern Australia. Beagle 15:1–21
Wehe T (2006) Revision of the scale worms (Polychaeta: Aphroditoidea) occurring in the seas surrounding the Arabian Peninsula. Part I: Polynoidae. Fauna Arabia 22:23–197
Williams C (2009) The secret weapon. http://www.champwilliams.com/limpet_high_res.html
Acknowledgments
We are grateful to Dr. Massimo Boyer for his underwater pictures and for the sample of Medioantenna variopinta sp. nov. collected in February 2008, to Dr. F. Falcieri for providing the map of the study area, to Dr. R. Barnich for fruitful discussions of worm taxonomy and to Prof. Imajima and Dr. H. Namikawa, Dr.W. Sato-Okoshi and Dr. E. Kuprianova for providing us the paratype of Medioantenna clavata. The study has been supported by the Russian Foundation for Basic Research (08-04-92244-GFEN-a and 09-05-00736a) and the Italian Ministry of Foreign Affairs (MAE) and is a contribution to the Consolidated Research Group of the “Generalitat de Catalunya” (ref. numb. 2009SRG665).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Martin Thiel.
Rights and permissions
About this article
Cite this article
Di Camillo, C.G., Martin, D. & Britayev, T.A. Symbiotic association between Solanderia secunda (Cnidaria, Hydrozoa, Solanderiidae) and Medioantenna variopinta sp. nov. (Annelida, Polychaeta, Polynoidae) from North Sulawesi (Indonesia). Helgol Mar Res 65, 495–511 (2011). https://doi.org/10.1007/s10152-010-0239-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10152-010-0239-7