- Original Article
- Published:
A review of the genus Mirostenella Bayer, 1988 (Octocorallia: Primnoidae) with a description of a new subgenus and species
Helgoland Marine Research volume 67, pages 229–240 (2013)
Abstract
In the present study, the genus Mirostenella Bayer (Proc Biol Soc Wash 101:251–256, 1988) is revised. We add to knowledge of the type species of the genus M. articulata Bayer (Proc Biol Soc Wash 101:251–256, 1988) from newly collected material from the Antarctic cruise Polarstern ANT XIX/5, and an illustrated description of this species is included. Cairns and Bayer (Smithson Contrib Zool 629:1–79, 2009) included Dicholaphis delicatula Thomson and Rennet (Sci Rept C Zool Bot 9(3):1–46, 1931) in the genus Mirostenella but after the examination of the type material, it is proposed to include the species in the recently described Plumarella subgenus, Faxiella (Zapata-Guardiola and López-González in Sci Mar 76:357–380, 2012). In addition, a new species of Plumarella, Plumarella castellviae sp. nov. from SubAntarctic waters is also described and illustrated. The species has similarities to Mirostenella but differs from it in the absence of organic nodes at bifurcation points and the presence of a sympodial branching pattern. Moreover, a new subgenus, Verticillata, is also proposed to include Plumarella species with polyps arranged in whorls around branchlets.
Introduction
The Primnoidae family has been studied, since it was described by Gray (1858), on numerous occasions by numerous authors (Wright and Studer 1889; Versluys 1906; Roule 1908; Gravier 1913; Kükenthal 1919; Deichman 1936; Broch 1965; Bayer 1998; López-González et al. 2002; Cairns 2006; inter alios). The major taxonomic contributions concerning this family are the reports of the Challenger Expedition (Wright and Studer 1889), the Siboga Expedition (Versluys 1906) and the Albatross Expeditions (Nutting 1908, 1909, 1912), as well as the publications of Kinoshita (1907, 1908a, b) on Japan primnoids and the revisions given by Kükenthal (1915, 1924). Doubtless, one of the most prolific authors on primnoids is Bayer (1955, 1988, 1998, among others) who, along with other authors like Cairns (2006), has made the family Primnoidae one of the most investigated gorgonian families.
Although recently a valuable generic review of family Primnoidae was carried out by Cairns and Bayer (2009), a more detailed revision of some genera is needed. One of the most unusual primnoid genera was that described by Bayer (1988) as Mirostenella. This monospecific genus has typical primnoid sclerites but the calcified axis includes organic nodes, and this remarkable feature was the most important aspect discussed by the original author. Cairns and Bayer (2009) cautiously placed a second species in the genus, Mirostenella delicatula (Thomson and Rennet, 1931), originally described in genus Dicholaphis Kinoshita, 1908 and also referred to an unpublished work on a possible third species of Mirostenella.
The main objective of this paper is the revision of the species currently included in the genus Mirostenella using newly collected material from recent Antarctic cruises as well as the type material and original descriptions.
Materials and methods
Type material of Dicholaphis delicatula Thomson and Rennet, 1931 is deposited in the Australian Museum (AM), which kindly sent us the specimens for light microscopy and scanning electron microscope (SEM) examination of colony shape, polyp morphology and sclerite variability.
The material here studied was collected on the R/V Polarstern cruises ANT XIII/4 (17 March–20 May 1996), ANT XIX/5 (LAMPOS, Latin American Polarstern Study, 3 April–5 May 2002), Discovery Expedition on board RSS William Scoresby (November 1931–April 1932), and from various USARP (United States Antarctic Research Program) and USAP (United States Antarctic Program) cruises on board the R/V Eltanin, R/V Hero and R/V Lawrence M. Gould (LMG).
The material of R/V Polarstern cruises was collected using an Agassiz trawl along Tierra del Fuego and South Georgia Islands; USARP and USAP material was from Tierra del Fuego, the Falkland Islands and Burdwood Bank (Fig. 1). Specimens collected on R/V Polarstern cruises were fixed in 10 % buffered formalin and then transferred to 70 % ethanol.

Distribution of the primnoid species here studied: circle, Mirostenella articulata; square, Plumarella delicatula; triangle, Plumarella castellviae sp. nov. Empty symbols represent the respective holotypes
Fragments from different parts of the colonies were prepared for study by SEM employing the usual methodology described by Bayer and Stefani (1988), and Alderslade (1998); permanent mounts were made for light microscopy. Visualization of polyps and coenenchymal sclerites was improved by staining with methylene blue. Most of the stubs coming from material of USARP and USAP expeditions were prepared by Frederick Bayer and deposited in the Smithsonian Institution.
The colony and sclerite terminology mainly follows the method described by Bayer et al. (1983). The material studied is deposited at the National Museum of Natural History, Smithsonian Institution, Washington, D.C. (USNM); at the Natural History Museum, London (NHMUK); at the Museu de Zoologia de Barcelona, Barcelona, Spain (MZB); and in the octocoral reference collection of the research group “Biodiversidad y Ecología de Invertebrados Marinos” of the University of Seville (BEIM-CRO).
Results
Diagnosis (modified from Bayer (1988) in bold)
Primnoidae with a planar colony shape that is dichotomously branched. Straight polyps arranged in pairs or whorls of 3–6, almost perpendicular or inclined upward towards branchlets, with 8 complete rows of body scales. Thorny marginal scales with smooth outer surface aligned with opercular scales, which do not fold over them. Calcified axis interrupted by organic nodes at points of bifurcation.
Type species:
Mirostenella articulata Bayer, 1988
-
Mirostenella articulata Bayer, 1988 (Figs. 2, 3, 4, 5)
Fig. 2 
Mirostenella articulata, BEIM(CRO-58): a, b two colonies
Fig. 3 
Mirostenella articulata, BEIM (CRO-58). Detail of a branchlet, stereo pair
Fig. 4 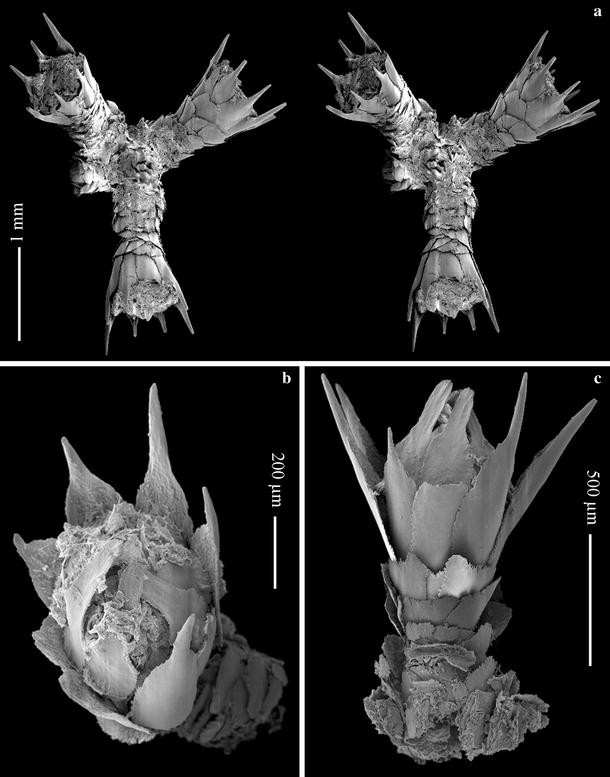
Mirostenella articulata, BEIM (CRO-58): a polyp whorl, adaxial view, stereo pair; b polyp, oral view; c polyp, adaxial view
Fig. 5 
Mirostenella articulata, BEIM (CRO-58): a opercular scales; b marginal scales; c body scales; d coenenchymal scales
-
Mirostenella articulata Bayer, 1988: 251–256—Cairns and Bayer 2009: 28 (listed), 38–39, fig. 10 r-a.




Examined material
Type material
USNM 79959, holotype, USAP, Eltanin 22, stn 1536, 54°28.98′S, 39°22.02′W, South Georgia Island, off west tip of island, Scotia Sea, SubAntarctic, 659–686 m depth, 8 February 1966, one colony; USNM 79960, two paratypes, same data as the holotype.
Additional material
BEIM (CRO-58), Polarstern cruise ANT XIX-5, stn PS61-164-01, 53°23.8′S, 42°42.03′W, Shag Rocks, west South Georgia, Antarctica, 312–322 m depth, 9 April 2002, two colonies, one of them with holdfast. MZB (2012-0484), Polarstern cruise ANT XIX-5, stn PS61-167-01, 53°23.68′S, 42°42.23′W, Shag Rocks, west South Georgia, Antarctica, 306–342.7 m depth, 9 April 2002, one fragment of colony.
Description of the additional material examined
Smaller colony (Fig. 2a) 4.5 cm tall and about 4 cm wide, with holdfast. Larger colony, fragmented (Fig. 2b), 10.5 × 8 cm. Fragment of colony 16 × 3 cm. Plane colonies dichotomously branched, internodes about 5–33 mm in length, terminal branchlets up to 55 mm. Axis ochre, organic nodes dark brown, more distinct in distalmost nodes. Basal axis diameter up to 1.2 mm and 0.9 cm long before the first bifurcation in the small colony.
Polyps straight, almost perpendicular or inclined upward towards branchlets, arranged in whorls (Fig. 3), 3–6 polyps per whorl and 4–6 whorls per cm. Polyps (Figs. 3, 4) funnel shape; about 1.0–2.0 mm in height and 0.48–0.7 mm in diameter. Polyp body with 8 longitudinal rows of scales, 5–6 scales in each abaxial row and 4 scales in each adaxial row; proximal scales disorganized and overlapping one another.
Eight opercular scales (Fig. 5a) generally isosceles triangle-shaped, 0.43–0.58 × 0.21–0.28 mm, although some sclerites are strongly reduced in width on the distal two-thirds, 0.09–0.13 mm in width distally. Inner surface tuberculated, crowded at least on the proximal half of scale, forming short longitudinal ridges distally, but keels are absent. Outer surface granulated. Basal margin tuberculated, free margin finely serrated.
Eight marginal scales, 0.64–1.09 × 0.23–0.32 mm in size (Fig. 5b), basal part oval shaped supporting a long projecting thorn that is round in cross-section, almost smooth, and occupies more than one-third of the total sclerite length. Inner surface of basal part of scale completely tuberculated, without a keel; outer surface almost completely smooth, with only a few tubercles on most proximal border. Basal and free margin with minute irregular processes.
Body scales (Fig. 5c) oval-fan shape, 0.17–0.39 × 0.23–0.40 mm. Inner surface tuberculated; outer surface granulated, proximal border with tubercles. Margin with irregular processes.
Coenenchymal sclerites (Fig. 5d) round, oval, rectangular-shaped, 0.14–0.45 mm in maximum length; inner surface tuberculated, outer surface granulated, free margin with irregular processes.
Geographical and bathymetrical distribution
At present, Mirostenella articulata is only known from South Georgia Island, Antarctica (Fig. 1) (Bayer 1988, Cairns and Bayer 2009, present study), between 201 and 210 m in depth (Cairns and Bayer 2009), 312–342.7 m in depth (present study) and 659–686 m in depth (Bayer 1988).
Remarks
A re-description of the holotype deposited in the USNM has not been necessary due to the complete original description given by Bayer (1988). The general colonial structure, polyp distribution and sclerites size and shape of the additional material examined are quite similar to that of the holotype. Currently, the knowledge of this species is very limited. Three colonies were used in the original description, a fourth colony was recorded by Cairns and Bayer (2009), and two more colonies and a fragment are here reported, all from the South Georgia area, between 201 and 686 m depth. Despite the intense sampling effort carried out by USARP, USAP and Polarstern expeditions, more records and information from other than usual Antarctic areas are needed to define a possible wider geographical distribution. Molecular analyses are also needed to facilitate discussion of the phylogenetic importance and implications of the distinct organic and non-organic nodes in octocoral families, such as primnoids and isidids.
-
Genus Plumarella Gray, 1870
Diagnosis (modified from Zapata-Guardiola and López-González (2012) in bold)
Primnoidae colonies usually uniplanar, alternately pinnately branched or dichotomously branched and may also be bottlebrush in shape. Polyps may occur in an alternate or opposite biserial arrangement, or in whorls on branchlets and also crowded on all sides of branchlets in no order. Polyps covered by 8 complete rows of body scales, adaxial rows sometimes reduced in number and size. Marginal scales arranged in one circlet of scales, which do not fold over the opercular scales, may have ridges, spines and other ornamentations in their inner surface but never will be keeled.
-
Plumarella (Faxiella) delicatula Thomson and Rennet, 1931 (Figs. 6, 7)
Fig. 6 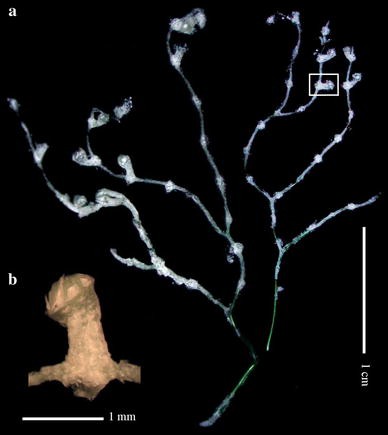
Plumarella (Faxiella) delicatula, holotype (AM G13266): a colony in two fragments; b detail of a polyp indicated in a, lateral view
Fig. 7 
Plumarella (Faxiella) delicatula, holotype (AM G13266): Scales. Note the fragmented state of the material examined. op Opercular scale; bs body scales; cs coenenchymal scale
-
Dicholaphis delicatula Thomson and Rennet, 1931: 30, pl. 9, fig. 8–9, pl. 12, fig. 4.
-
Mirostenella delicatula, Cairns and Bayer 2009: 28 (listed), 38–39.


Examined material
AM (G13266), holotype, Australasian Antarctic Expedition 1914, 54°36′S, 158°48′E, off Macquarie Island, SubAntarctic, 2,743 m depth.
Remarks
The current state of conservation of the type material is very poor for the comparative taxonomic study here proposed. The colony is partially decalcified, and the few polyps and sclerites present are often fractured. Without enough material to work on, no SEM analysis could be carried out of polyps and branchlets; only one or two polyps were dissolved and analysed by SEM to observe the sclerite shape and ornamentation.
Description of the holotype
Two delicate uniplanar fragments (Fig. 6a), 2.6 and 3.3 cm tall and about 1 and 1.5 cm wide, respectively, dichotomously branched, length between two bifurcation points about 4–7 mm, terminal branchlets up to 18 mm.
Polyps cylindrical, about 1.1–1.8 mm long and 0.37–0.69 mm in diameter (Fig. 6b), curved upward towards the branchlets and placed in pairs, 4 pairs per cm (Fig. 6a).
Outer surface of polyp scales smooth (Fig. 7), inner surface tuberculated. Free margin smooth. Marginal scales about 0.52 × 0.3 mm, body scales (Fig. 7bs) about 0.16–0.28 mm in maximum diameter and coenenchymal scales (Fig. 7cs) about 0.09 mm in maximum diameter.
Geographical and bathymetrical distribution
At present, Plumarella (Faxiella) delicatula is known from off Macquarie Island (Fig. 1), SubAntarctic, and 2,743 m in depth.
Remarks
The material here examined is the only type material available for Plumarella (Faxiella) delicatula. This specimens are very fragile and delicate, with only two fragments, and just a few badly preserved polyps, remaining of the original colony described by Thomson and Rennet (1931: pl. 9, fig. 8)
Thomson and Rennet placed the specimen in Kinoshita’s genus Dicholaphis due to the delicacy of the colony, its slender axis, the dichotomous branching pattern in one plane, and polyps, with a high operculum of 8 scales, curved towards the branchlets.
Cairns and Bayer (2009: 38) proposed, without further comments, that Dicholaphis delicatula should be included in Mirostenella as a new combination. However, they did not have the opportunity to examine the type specimen (S. Cairns pers. comm.) and not being completely sure of their decision, they listed the new combination with a question mark (Cairns and Bayer 2009: 28).
After examination of the type specimen, all characters used by Thomson and Rennet to place this specimen in Dicholaphis have been confirmed (except for the number of scales on the polyps due to many scales being missing on the scarce polyps), and considering organic nodes that are found only in Mirostenella, the genera are easily differentiated on this character. Due the state of preservation of the examined material, and according to the original texts available and the present examination, we think that it would have been preferable not place the species into the genus Mirostenella (Cairns and Bayer 2009).
Formerly genus Dicholaphis included only the type species, Dicholaphis delicata Kinoshita, 1908 which differs from Dicholaphis delicatula Thomson and Rennet, 1931 by the arrangement of polyps (singly placed in the Japanese species, and in pairs in the SubAntarctic species).
Nevertheless, the genus Dicholaphis has been recently incorporated as a subgenus of Plumarella (Cairns 2011) due to their similarities but being otherwise distinguished by the singly arrangement of polyps on all sides of the branchlets, and leaving in Plumarella (Plumarella) those species with an alternate biserial polyp arrangement.
In the last decade, the increasing number of taxonomic contributions in the study of primnoid gorgonians has led to the description of numerous new species and genera, each time in agreement with the current generic diagnostic criteria. Unfortunately, an increase in the number of taxa and general knowledge of the morphologic diversity in the primnoidae has resulted in a relative instability and inconsistency of the current systematics at generic level (having names changing from a generic to subgeneric status, and viceversa). On the one hand, some characters are used to distinguish genera (e.g. arrangement of polyps on branchlets); and, on the other hand, they are also used into differentiate subgenera in highly diversified, speciose genera such as Thouarella, and more recently in Plumarella. In any case, we should describe the observed variability and assign a taxonomic level of importance, although it could be tentative or provisional due the rate of new contributions being published in this family with the discovery of new morphological forms. In this unexpected scenario, due to the number of taxonomical novelties being described in such a short time-scale [in comparison with other taxonomic groups, even within the octocorals, where we need to wait decades for new records or new taxa (mainly due by the lack of available specialist or a special political and scientific framework like the International Polar Year was)], we tentatively agree with Cairns to consider the character “arrangement of polyps on branchlets” at the subgeneric level.
According to this new wider definition of genus Plumarella including different polyp arrangements, the species of Thomson and Rennet should be included in this genus. However, the arrangement of polyps on branchlets, which is the character used to differentiate both subgenera (Plumarella and Dicholaphis), is in a biserial opposite arrangement in Plumarella delicatula and thus it should be included in the recently described subgenus, Faxiella (Zapata-Guardiola and López-González 2012). Plumarella abietina is the only species included in subgenus Faxiella and mainly differs from Plumarella delicatula in having larger polyps (up to twice the size) resulting in less pair of polyps per centimetre (a couple in P. abietina and four in P. delicatula).
-
Subgenus Verticillata subgen. nov.
Diagnosis
Plumarella in which funnel-shaped polyps occur straight and almost perpendicular to stem, placed in whorls along branchlets.
Etymology
From the modern Latin adjective verticillatus that means “disposed in or forming whorls” which comes from the Latin noun verticillus “whorl,” in reference to the arrangement of polyps in whorls on branchlets.
-
Gender: feminine.
Type species
Plumarella (Verticillata) castellviae sp. nov. here designated.
-
Plumarella ( Verticillata ) castellviae sp. nov. (Figs. 8, 9, 10, 11)
Fig. 8 
Plumarella (Verticillata) castellviae sp. nov., holotype (USNM 1099626): a whole colony; b detail of a branchlet
Fig. 9 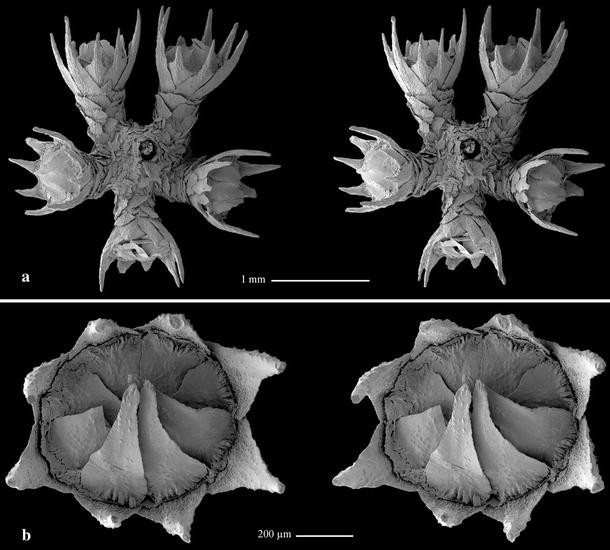
Plumarella (Verticillata) castellviae sp. nov., holotype (USNM 1099626): a polyp whorl, adaxial view, stereo pair; b polyp, oral view, stereo pair
Fig. 10 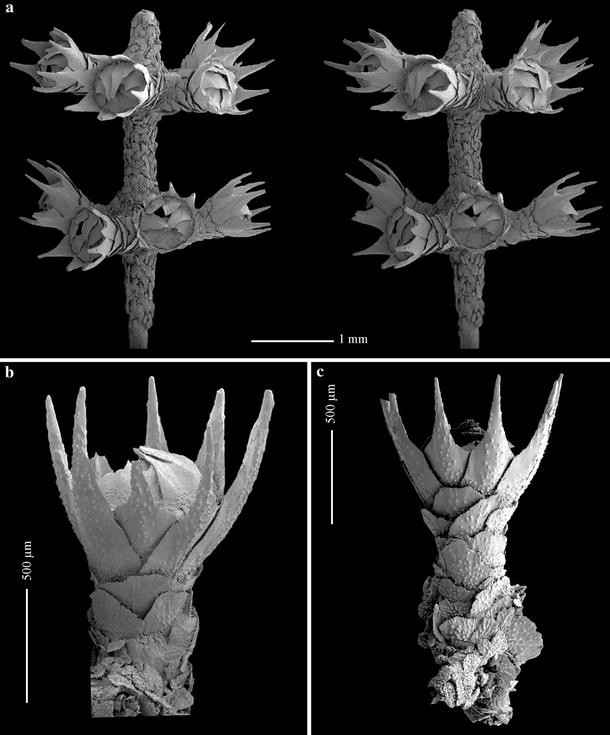
Plumarella (Verticillata) castellviae sp. nov., holotype (USNM 1099626): a polyp whorls, stereo pair; b polyp, adaxial view; c polyp, abaxial view
Fig. 11 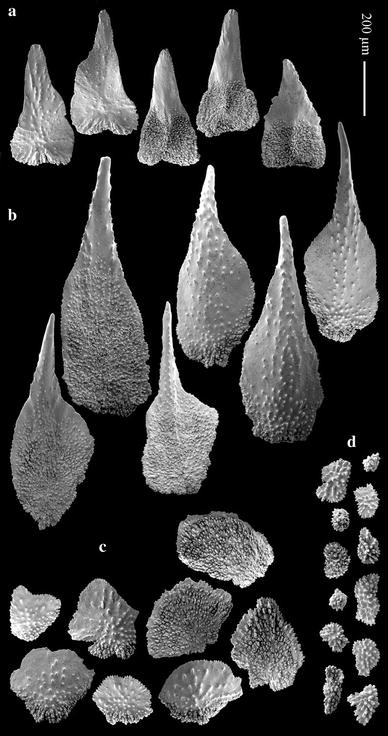
Plumarella (Verticillata) castellviae sp. nov., holotype (USNM 1099626): a opercular scales; b marginal scales, c body scales; d coenenchymal scales




Examined material
Type material Holotype: USNM 1099626, LMG 06-05, stn 2, 53°48′S, 64°54′W, off Isla Grande, Tierra del Fuego, SubAntarctic, 120 m depth, 15 May 2006, one dry colony. Paratypes: USNM 1128562, same data as holotype, nine fragmented colonies; NHMUK 2011.41, Discovery Expeditions, RSS William Scoresby, stn WS 877, 52°35.4′S, 61°3.6′W, south west of Falkland Islands, SubAntarctic, 350 m depth, 4 April 1932, two fragments; USNM 100787 (SEM stubs 1609, 1610), USNM 100795 and USNM 1019337, USAP, Eltanin 6, stn 339, 53°07.9′S, 59°24′W, west of Beauchene Island, Falkland Islands, SubAntarctic, 512–586 m depth, 3 December 1962, 1, 3 and 20 fragmented colonies respectively; USNM 58167 (plus SEM stubs 285, 297, 298) and USNM 88822 (plus SEM stub 2236), USAP, Eltanin 9, stn 740, 56°06′S, 66°18′W, east of Cape Horn, Drake Passage, Tierra del Fuego, SubAntarctic, 384–494 m depth, 18 September 1963, 10 and 5 colonies respectively; USNM 77392 (plus SEM stubs 1568, 1569), USAP, Eltanin 22, stn 1592, 54°45′S, 55°37′W, Burdwood Bank, Scotia Sea, SubAntarctic, 1,647–2,044 m depth, 14 March 1966, ten fragmented colonies; USNM 100789 (plus SEM stubs 1591, 1592), USAP, Hero 715, stn 885, 54°55′S, 64°9.7′W, Staten Island, Tierra del Fuego, SubAntarctic, 493–511 m depth, 30 October 1971 one colony; BEIM (CRO-57), Polarstern cruise ANT XIII/4, stn 40/111, 55°28.8′S, 66°04.5′W, southeast of Isla Nueva, Tierra del Fuego, SubAntarctic, 1,147 m depth, 17 May 1996, one fragment; MZB (2012-0485), Polarstern cruise ANT XIII/4, stn 40/115, 55°27.4′S, 66°05.7′W, southeast of Isla Nueva, Tierra del Fuego, SubAntarctic, 580 m depth, 18 May 1996, numerous fragments; MZB (2012-0486), Polarstern cruise ANT XIX/5, stn PS61/150-1, 54°30.22′S, 56°8.20′W, south east of Falkland Islands, Antarctica, 286.3–290.3 m depth, 6 April 2002, two fragmented colonies; MZB (2012-0487), Polarstern cruise ANT XIX/5, stn PS61/153-1, 54°31.22′S, 56°8.93′W, south east of Falkland Islands, Antarctica, 277.2–295.8 m depth, 6 April 2002, one colony.
Description of the holotype
Uniplanar colony (Fig. 8) 25 cm tall and about 20 cm wide, moderate sympodial branching with branchlets up to 6 cm in length, simple or branched up to the sixth order. Axis brown, broken in its proximal portion, without holdfast. Axial nodes without reduction in calcification. Basal axis diameter about 3 mm.
Polyps funnel-shaped, straight, about 0.9–1.4 mm in height and 0.37–0.50 mm in diameter, almost perpendicular to branchlet and placed in whorls (Fig. 8b); 5–7 polyps per whorl (Fig. 9a) and 6–8 whorls per cm (Fig. 8b). Polyp body with 8 longitudinal rows of scales overlapping one another, 3 scales in each adaxial row (Fig. 10b), 5–6 scales in each abaxial row but disorganized proximally (Fig. 10c).
Operculum low with 8 triangle-shaped scales (Fig. 9b, 11a) 0.40–0.47 × 0.19–0.26 mm in size. Proximal half of inner surface tuberculated, distal half smooth, without keel. Outer surface radially granulated. Basal margin tuberculated, free margin finely serrated almost smooth.
Eight marginal scales (Fig. 11b), 0.67–0.99 × 0.26–0.32 mm, spoon-shaped with distal thorn. Inner surface tuberculated until thorn base; pointed granules on thorn surface. Outer surface with pointed granules arranged longitudinally, proximal part tuberculated. Free margin finely serrated, basal margin tuberculated.
Body scales (Fig. 11c) oval-fan shaped, 0.18–0.37 × 0.21–0.37 mm. Inner surface almost completely tuberculated. Proximal outer surface tuberculated, distal outer surface granulated, basal outer margin tuberculated, free margin finely serrated.
Coenenchymal scales (Fig. 11d) irregular oval shaped, 0.06–0.24 mm in maximum length; inner surface tuberculated, outer surface granulated.
Variations from holotype
The colony shape varies from uniplanar to a bushy appearance. Their size range is between 11 and 25 cm tall and 6–20 cm wide. Polyps are arranged in whorls from 4 to 7, and their density ranges from 5 to 8 whorls per centimetre. The funnel form of the polyps is similar to those of the holotype, from 0.8 to 1.6 mm long and from 0.3 to 0.5 mm in diameter. The distribution and form of the sclerites from polyps and coenenchyme are as in the holotype.
Geographical and bathymetrical distribution
At present, Plumarella castellviae sp. nov, is known from SubAntarctic waters (Fig. 1), from Tierra del Fuego and the Falkland Islands to Burdwood Bank in the Scotia Sea, between 120 and 2,044 m in depth.
Etymology
The species is named after Dr. Josefina Castellví (Institute of Marine Sciences, Spain) in recognition of her major contribution to the understanding of Antarctic ecosystems and, such as the first manager of the Spanish Antarctic Program, for being a key person for the Spanish Antarctic research.
-
Gender: feminine.
Remarks
Plumarella (Verticillata) castellviae sp. nov. is characterized by having polyps arranged in whorls. This character together with the presence of eight un-keeled marginal scales aligned with the operculars indicates a close relationship to the genus Mirostenella. However, it can be easily distinguished from Mirostenella by not having its calcified axis interrupted by organic nodes and also by having an unusual colonial branching pattern among primnoids, a profusely branched sympodial form that resembles the “loosely alternate pinnate, branches slightly geniculate at points of branchlet origin” form described for Plumarella circumoperculum Cairns 2010 (see Fig. 1C in Cairns 2010). This branching pattern together with the lack of keel on marginal scales place the new species within the genus Plumarella. The subgenera included in the genus are distinguished by the arrangement of polyps on branchlets; however, none of them include polyps in whorls, which is the state of the character in Plumarella castellviae; therefore, a new subgenus Verticillata has been designated to include it.
Nonetheless, future contributions with more information (including different source of data and the description of new taxa currently unknown) should surely depict a more natural (and less artificial) systematics for the Family Primnoidae and evaluate the suitability of the available generic and subgeneric names.
References
Alderslade P (1998) Revisionary systematics in the gorgonian family Isididae, with description of numerous new taxa (Coelenterata, Octocorallia). Rec W Aust Mus 55:1–359
Bayer FM (1955) Contributions to the nomenclature, systematics, and morphology of the Octocorallia. Proc US Nat Mus 105(3357):207–220
Bayer FM (1981) Key to the genera of Octocorallia exclusive of pennatulacea (Coelenterata: Anthozoa), with diagnoses of new taxa. Proc Biol Soc Wash 94(3):902–947
Bayer FM (1988) Mirostenella articulata, a remarkable new genus and species of Primnoid Octocoral (Gorgonacea) with uncalcified axial nodes. Proc Biol Soc Wash 101(2):251–256
Bayer FM (1998) A review of the circumaustral gorgonacean genus Fannyella Gray, 1870 with description of five new species (Coelenterata: Octocorallia: Primnoidae). Senckenb Biol 77:161–204
Bayer FM, Stefani J (1988) Primnoidae (Gorgonacea) de Nouvelle-Calédonie. Bull Mus Natn Hist Nat Paris 10(A)3:449–476
Bayer FM, Grasshoff M, Verseveldt J (1983) Illustrated trilingual glossary of morphological and anatomical terms applied to Octocorallia. Brill, Leiden, pp 1–75
Broch H (1965) Some octocorals from Antarctic Waters. Scientific Results of the “Brategg” Expedition, 1947–48. Christensens Hvalfangstmus i Sandejford Publ 26:19–38, 1–7
Cairns SD (2006) Studies on western Atlantic Octocorallia (Coelenterata: Anthozoa). Part 6: the genera Primnoella Gray, 1858; Thouarella Gray, 1870; Dasystenella Versluys, 1906. Proc Biol Soc Wash 119(2):161–194
Cairns SD (2010) Review of Octocorallia (Cnidaria: Anthozoa) from Hawaii and Adjacent Seamounts. Part 3: genera Thouarella, Plumarella, Callogorgia, Fanellia, and Parastenella. Pac Sci 64(3):413–440
Cairns SD (2011) A revision of the Primnoidae (Octocorallia: Alcyonacea) from the Aleutian Islands and Bering Sea. Smithson Contrib Zool 634:1–55
Cairns SD, Bayer FM (1954) Studies on western Atlantic Octocorallia (Coelenterata: Anthozoa). Part5: the genera Plumarella Gray, 1870; Acanthoprimnoa, n. gen.; and Candidella Bayer, 1954. Proc Biol Soc Wash 117(4):447–487
Cairns SD, Bayer FM (2009) A generic revision and phylogenetic analysis of the Primnoidae (Cnidaria: Octocorallia). Smithson Contrib Zool 629:1–79
Deichman E (1936) The Alcyonaria of the western part of the Atlantic Ocean. Mem Mus Comp Zool Harvard 53(1–317):1–37
Gravier CH (1913) Seconde Expédition Antarctique Française (1908–1910). Alcyonaires (2° note préliminaire). Bull Mus Nat Hist Nat Paris 19:589–593
Gray JE (1858) Synopsis of the families and Genera of Axiferous zoophytes or barked corals. Proc Zool Soc Lond 1857:278–294
Kinoshita K (1907) Vorläufe Mitteilung uber Einige Neue Japanische Primnoidkorallen. Annot Zool Jap 6(3):229–237
Kinoshita K (1908a) Primnoidae von Japan. J Coll Sci Imp Univ Tokyo 23(12):1–74
Kinoshita K (1908b) Diplocalyptra, eine neue Untergattung von Thouarella (Primnoidae). Annot Zool Jap 7(1):49–60
Kükenthal W (1915) System und Stammesgeshichte der Primnoidae. Zool Anz 46(5):142–158
Kükenthal W (1919) Gorgonaria. Wissensch Ergebn deutschen Tiefsee-Exped ‘‘Valdivia’’ 13(2):1–946, 30–89
Kükenthal W (1924) Coelenterata: Gorgonaria Das Tierreich 47. Walter de Gruyter and Co, Berlin, pp 1–478
López-González PJ, Gili JM, Orejas C (2002) A new primnoid genus (Anthozoa Octocorallia) from the Southern Ocean. Sci Mar 66:383–397
Nutting CC (1908) Descriptions of the Alcyonaria collected by the U.S. Bureau of Fisheries Streamer Albatross in the Vicinity of the Hawaiian Islands in 1902. Proc US Nat Mus 34:543–601
Nutting CC (1909) Alcyonaria of the Californian coast. Proc US Nat Mus 35:681–727
Nutting CC (1912) Descriptions of the Alcyonaria collected by the U.S. Bureau of Fisheries Streamer Albatross, mainly in Japanese waters during 1906. Proc US Nat Mus 43:1–104
Roule (1908) Alcyonaires. Expédition Antarctique Française (1903–1905). Sci Nat Doc Sci 15:1–6, pl 1
Thomson JA, Rennet N (1931) Alcyonaria Madreporaria and Antipatharia Australasian Antarctic Exped. Sci Rept (C- Zoology and Botany) 9(3):1–46
Versluys J (1906) Die Gorgoniden der Siboga-Expedition. II. Die Primnoidae. Siboga-Expeditie 13a:1–187
Wright EP, Studer T (1889) Report on the Alcyonaria collected by H.M.S. Challenger during the years 1873–1876. Rep Sci Res Challenger 31:i–1xxvii, 1–314
Zapata-Guardiola R, López-González PJ (2012) Revision and redescription of the species previously included in the genus Amphilaphis Studer and Wright in Studer, 1887 (Octocorallia: Primnoidae). Sci Mar 76(2):357–380
Acknowledgments
The authors would like to thank Stephen Cairns (USNM) for his assistance and comments on the family Primnoidae during a long stay in the National Museum of Natural History and for allowing us access to all Bayer’s material. We also acknowledge Emma Sherlock and the Zoology Department for access to the collection of NHMUK, and to Dr. Stephen Keable (Australian Museum) for lending, the type of Plumarella delicatula for examination. We are grateful to the officers and crew of the R/V Polarstern and many colleagues on board during the LAMPOS cruise for their support and help and also to Erika Mutschke and Carlos Rios (Universidad de Magallanes, Chile) for making available the material collected during the Polarstern cruise ANT XIII/4. We would also thank the editor of Helgoland Marine Research and give special thanks to the referees for their ideas and comments about the manuscript, which significantly improved it. Support for this study was provided by the Spanish CICYT projects—CLIMANT—POL2006-06399/CGL (ANT XXIII/8) and REN2001-4929-E/ANT (LAMPOS). Mr. Tony Krupa is thanked for reviewing the English version.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by P. Funch.
Rights and permissions
About this article
Cite this article
Zapata-Guardiola, R., López-González, P.J. & Gili, JM. A review of the genus Mirostenella Bayer, 1988 (Octocorallia: Primnoidae) with a description of a new subgenus and species. Helgol Mar Res 67, 229–240 (2013). https://doi.org/10.1007/s10152-012-0318-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10152-012-0318-z