- Original Article
- Published:
Description of two species of Caprella (Crustacea: Amphipoda: Caprellidae) from the North Pacific; C. californica Stimpson, 1857 and C. scauroides Mayer, 1903, with a new appraisal of species ranking for C. scauroides
Helgoland Marine Research volume 67, pages 371–381 (2013)
Abstract
Caprella californica Stimpson, 1857 [sensu lato] is one of the dominant species of Caprella spp. (Crustacea: Amphipoda) along the Pacific coast of Mexico to Canada and Japan to Hong Kong, China, and recently widely reported from the Australian coasts. Detailed morphological comparison of C. californica [sensu lato] collected from California and the Uwa Sea, Japan, revealed several diagnostic differences, including: (1) the body somites of the former are more slender than those of the latter; (2) in the male of the former, head possesses an anteriorly curved dorsal projection, while in the latter, head possesses a straight, forward-pointing dorsal projection; (3) in the former, pereonite 5 is longer than pereonites 3 and 4, while in the latter, pereonite 5 is the same length as pereonite 3; (4) in the male of the former, propodus of gnathopod 2 has a small apical rectangular projection, while in the latter, the corresponding margin has a large round projection; and (5) in the former, pereopod 7 is slender with the merus and propodus subequal, while pereopod 7 in the latter is robust with the merus shorter than propodus. These differences are indications of species level differences. Thus, we propose Caprella scauroides Mayer, 1903 to a species level for the Japanese C. californica [sensu lato]. A detailed description with illustrations of C. californica [sensu stricto] and C. scauroides is provided.
Introduction
Caprella californica Stimpson, 1857 [sensu lato] was reported from California during the 19th Century as one of the earlier reported species of the genus Caprella (Stimpson 1857). Mayer (1890, 1903) recognized 6 “formae” (=varieties), that is, Caprella scaura f. californica Stimpson from California, C. scaura f. cornuta Mayer, 1890 from Brazil, C. scaura f. diceros Mayer, 1890 from Japan, C. scaura f. scauroides Mayer, 1903 from Hong Kong, China and Japan, C. scaura f. spinirostris Mayer, 1903 from Chile, and C. scaura f. typica Mayer, 1890 from Brazil, under Caprella scaura Templeton, 1836 originally reported from Mauritius in the south Indian Ocean. According to recent rules of nomenclature, these “formae” are now treated as subspecies, and the nominal subspecies of C. scaura from Mauritius, the type locality, should be regarded as Caprella scaura scaura Templeton (see Krapp et al. 2006). Utinomi (1947) described C. scaura hamata Utinomi, 1947 as the 7th subspecies of C. scaura from Japan. Dougherty and Steinberg (1953) revived C. californica as a valid species, and McCain and Steinberg (1970) synonymized C. scaura scauroides from the western North Pacific and C. scaura spinirostris from Chile with C. californica [sensu lato]. Laubitz (1970) and Arimoto (1976) followed McCain and Steinberg (1970)’s nomenclature. Thus, C. californica [sensu lato] is recognized as widely distributed from the northern and southeastern Pacific, that is, British Colombia to the Pacific coast of Mexico (e.g., McCain and Steinberg 1970; Laubitz 1970; Watling and Carlton 2007), Chile (e.g., McCain and Steinberg 1970), and Japan to Hong Kong, China (e.g., McCain and Steinberg 1970; Arimoto 1976, 1978; Arimoto and Kikuchi 1977; Takeuchi 1999; Aoki and Takeda 2006; Lee and Hong 2009). Moreover, recently, C. californica [sensu lato] was widely reported from the Australian coast (Montelli 2010). However, Krapp et al. (2006) suggested the presence of 7 subspecies of C. scaura including Caprella scaura californica, Caprella scaura scauroides, and Caprella scaura spinirostris.
Nevertheless, detailed comparisons of these C. californica and C. scaura complexes have not been reported. Thus, as a first step toward clarification of these complexes, we conducted detailed taxonomic descriptions for Caprella californica [sensu lato] deposited at the California Academy of Sciences, California, USA, and those newly collected from the Uwa Sea, southwestern Japan. These described specimens are mature individuals, which possess all somite appendages. The comparison revealed that there are clear differences at the species level. Thus, C. scauroides Mayer, 1903 is proposed to be elevated from subspecies to species level, and detailed descriptions of C. californica Stimpson, 1857 [sensu stricto] and C. scauroides are provided.
Materials examined are deposited in the Australian Museum, New South Wales, Australia, the California Academy of Sciences, California, and the Museum of Comparative Zoology, Harvard University, Massachusetts, USA. Abbreviations used in the present study are as follows: AM, the Australian Museum; CASIZ, the California Academy of Sciences, Invertebrate Zoology; MCZ, the Museum of Comparative Zoology, Harvard University; A, antenna; ABD, abdomen; G, gnathopod; L, left; LL, lower lip; MD, mandible; MX, maxilla; MXP, maxilliped; P, pereopod; R, right; UL, upper lip.
-
Caprella californica Stimpson, 1857 (Figs. 1 , 2 , 3 , 4 )
Fig. 1 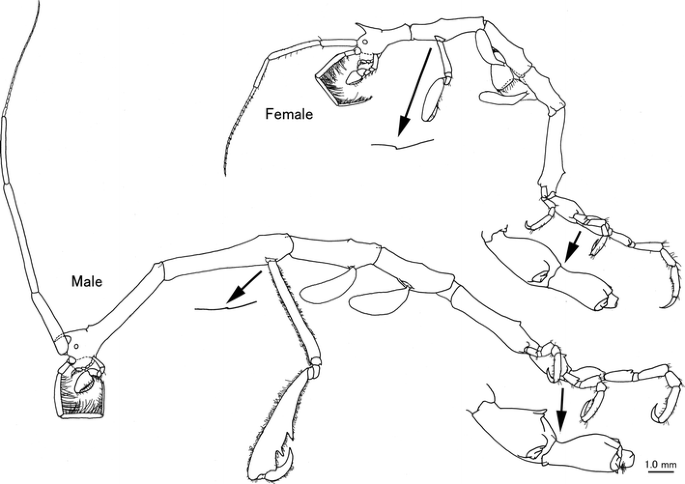
Caprella californica Stimpson, 1857 from San Diego, San Diego County, California, USA. Male “a”, CASIZ 189983, 24.23 mm; female “b”, CASIZ 189984, 15.55 mm
Fig. 2 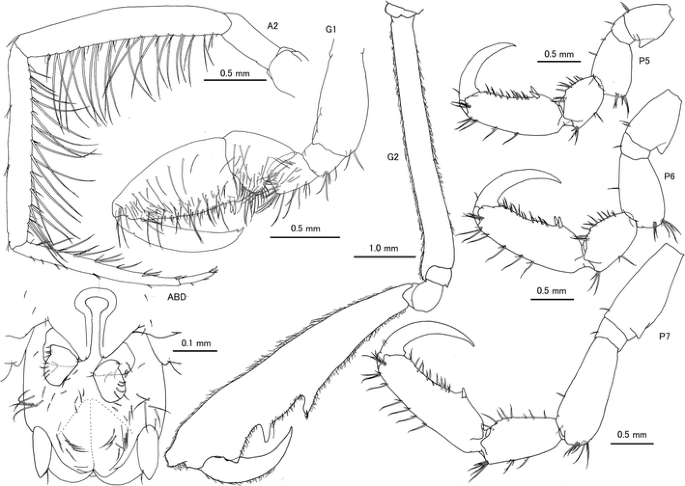
Caprella californica Stimpson, 1857 from San Diego, San Diego County, California, USA. Male “a”, CASIZ 189983, 24.23 mm, A2, G1, G2, P5, P6 and P7; male “e”, CASIZ 189985, 26.39 mm, ABD
Fig. 3 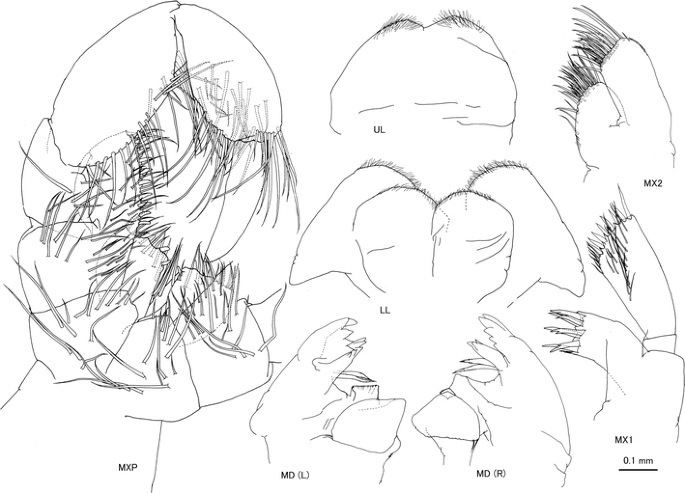
Caprella californica Stimpson, 1857 from San Diego, San Diego County, California, USA. Male “a”, CASIZ 189983, 24.23 mm
Fig. 4 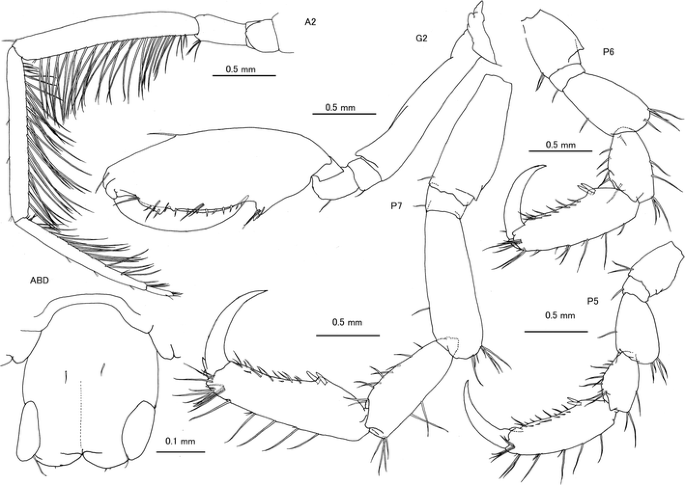
Caprella californica Stimpson, 1857 from San Diego, San Diego County, California, USA. Female “b”, CASIZ 189984, 15.55 mm, A2, G1, G2, P5, P6 and P7; female “f”, CASIZ 189986, 14.90 mm, ABD
-
Caprella californica Stimpson, 1857: 513–514; Boeck 1872, 35–37, 48, fig 1; Stimpson 1873, 97; Mayer 1882, 68–69; Dougherty and Steinberg 1953 44, 47; McCain and Steinberg 1970, 14 (in part); Laubitz 1970, 49–53, map fig 5b, fig 15; Martin 1977, 158–159; Watling and Carlton 2007, 624, 627, pl 309a.
Fig. 5 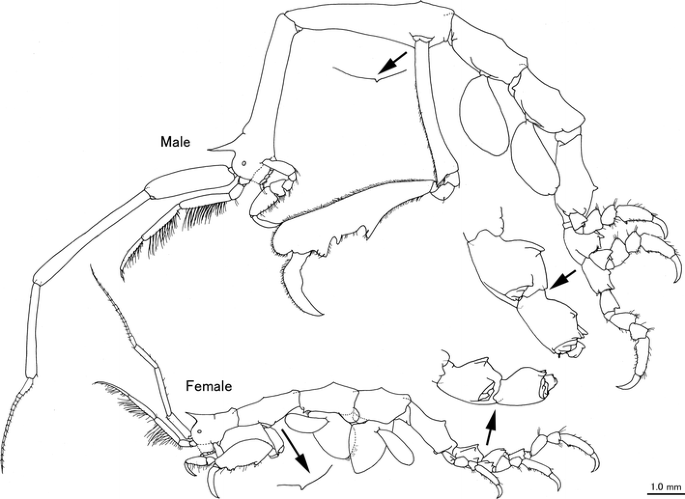
Caprella scauroides Mayer, 1903 from the Uwa Sea, southwestern Japan. Male “c”, AM-P.89082, 18.36 mm; female “d”, AM-P.89083, 9.49 mm
-
Caprella scaura [not Templeton, 1836]: Mayer 1890, 70–73 (in part).
-
Caprella scaura f. californica: Mayer 1903, 118–119, pl 5 fig 13.
-
Caprella scaura californica: Krapp et al. 2006, 3.





Material examined
Male “a”, CASIZ 189983, San Diego, San Diego County, California, USA, 16 June 1995, Coll. E.C. Starns; mature female “b”, CASIZ 189984, San Diego, San Diego County, California, USA, 16 June 1995, Coll. E.C. Starns; male “e”, CASIZ 189985, San Diego, San Diego County, California, USA, 16 June 1995, Coll. E.C. Starns; mature female “f”, CASIZ 189986, San Diego, San Diego County, California, USA, 16 June 1995, Coll. E.C. Starns; ca. 15 individuals, CASIZ 189987, San Diego, San Diego County, California, USA, 16 June 1995, Coll. E.C. Starns; 1 male, AM-P.89080 (ex. CASIZ 9785), San Diego, San Diego County, California, USA, 16 June 1995, Coll. E.C. Starns; 1 mature female, AM-P.89081 (ex. CASIZ 9785), San Diego, San Diego County, California, USA, 16 June 1995, Coll. E.C. Starns; 2 males, CASIZ 189988, Hog Island, Tomales Bay, Marin County, California, USA, Mudflat E of Hog Island, 2 April 1971, coll. Schneebeli W, Hansen R and Iverson E.
Description
Male “a”, CASIZ 189983. Body slender, 24.23 mm in body length. Pereonite 2 longest followed by pereonite 1 and pereonite 5. Head, 1.16 mm and pereonite 1, 4.09 mm; head and pereonite 1 partially fused; head with an anteriorly curved dorsal projection; eye, distinctive; pereonite 1 with a small anterodorsal projection and a dorsodistal projection. Pereonite 2, 5.26 mm with a small dorsal projection from 1/4 of posterior end and a minute ventral projection between gnathopod 2; height 0.25 × length. Pereonite 3, 3.67 mm with a small mid-dorsal projection and a dorsodistal projection. Pereonite 4, 3.16 mm with a small mid-dorsal projection and dorsolateral projection. Pereonite 5, 4.09 mm with a mid-dorsal projection. Pereonite 6, 1.58 mm with a pair of mid-dorsal projections. Pereonite 7, 1.21 mm with a pair of dorsodistal projections.
Antenna
1, 0.65 × body length; peduncular article 2 longest, 1.7 × article 1; article 3 subequal with article 1; flagellum with 20 articles, proximal article composed of 3 articles. Antenna 2, 0.4 × antenna 1; peduncular article 2 to flagellum article 1 with swimming setae.
Upper lip notched, wider than long, forming a pair of rounded projections, and finely setose. Lower lip well developed. Mandible left incisor with 5 teeth followed by lacina mobilis with 5 teeth and 2 accessory setal rows; molar well developed with a large flake-like projection and a small seta. Mandible right incisor with 5 teeth followed by lacina mobilis with 3 teeth and 2 accessory setal rows; molar well developed with a small molar flake and a small seta. Maxilla 1 outer plate with 7 stout apical setal-teeth; palp biarticulate; article 2, 5 × article 1 with a row of slender to robust marginal setae and 2 rows of lateral setae. Maxilla 2 inner plate oval with 2 rows of apical setae; outer plate with 2 rows of apical setae. Maxilliped lateral to basal margin densely setose; basal endite (inner plate) round with 3 stout teeth and a row of lateral setae; ischial endite (outer plate) subequal to inner plate with a row of apical setae; palp 4 articulate; article 2 longest, setose on inner margin; article 3, 0.8 × article 4, setose on lateral to apical margin; palp article 4 (dactylus) falcate.
Pereon
Gnathopod 1; basis subequal to propodus with 2 posterodistal setae; merus to carpus setose; propodus subtriangular, 2 × width, and densely setose on palm and lateral margin; palm begins near posterior margin with a pair of strong proximal spines; dactylus slightly curved distally.
Gnathopod 2 begins 1/8 along posterior margin of pereonite 2; basis subequal to pereonite 2, with dense fine setae on lateral parts and an anterodistal triangular projection; propodus subequal with basis and 4 × width; dorsal surface slightly convex and finely setose with a small rectangular apical projection; palm finely setose; proximal projection with a robust seta from 3/5 of proximal end; mid-palmar projection followed by a sinus and a small triangular projection; dactylus falcate, finely setose on lateral part.
Gill 3 oval, 2 × width and 0.75 × pereonite 3. Gill 4 similar to gill 3.
Pereopods 5 to 7 slender; pereopod 5, basis with a distal triangular projection; merus 1.5 × basis and 0.5 × propodus, laterally expanded with several setae; carpus subequal to basis; propodus 2 × basis, with a pair of stout proximal setae and ca. 10 pair of setae on palm; dactylus falcate. Pereopod 6 longer, 1.3 × pereopod 5. Pereopod 7 elongated, especially, basis, merus and carpus, 1.5 × pereopod 6 and 2 × pereopod 5; merus 1.2 × basis and subequal to propodus.
Pleon [Based on male “e”, CASIZ 189985, 26.39 mm]
Uropod 1 uniramus with 4 lateral setae. Uropod 2 uniramus with an apical seta. Telson round.
Female “b”, CASIZ 189984
Body length, 15.55 mm. Pereonite 5 longest followed by pereonite 2. Head, 1.03 mm and pereonite 1, 1.13 mm; head with an anterior forward-pointing dorsal projection; eye distinctive; pereonite 1 with a small dorsodistal projection; pereonite 2, 2.94 mm with a small mid-dorsal projection and a minute ventral projection between gnathopod 2; pereonite 3, 2.52 mm with a mid-dorsal projection; pereonite 4, 2.06 mm with a mid-dorsal projection and a minute dorsodistal projection; pereonite 5, 3.42 mm, with a mid-dorsal projection; pereonite 6, 1.42 mm; pereonite 7, 1.03 mm. Antenna 1 0.6 × body length; peduncular article 2 longest; flagellum with 19 articles, proximal article composed of 3 articles. Antenna 2 slender, 0.6 × antenna 1.
Pereon
Gnathopod 2 begins 2/5 along anterior margin of pereonite 2; basis 2/3 × pereonite 2 with an anterodistal triangular projection; carpus triangular; propodus subequal with basis and 2.5 × width; dorsal surface convex with 2 setae; palm scarcely setose; proximal projection with a robust seta from 2/5 of proximal end; mid-palmar projection followed by a minute sinus and a small triangular projection; dactylus falcate.
Pleon [female “f”, CASIZ 189986, 14.90 mm]
A seta instead of Uropod 1. Uropod 2 with an apical seta.
Distribution
Type locality: California, USA.
Other records: British Columbia, Canada; Friday Harbor, Washington Sate, USA; Magdalena Bay and Cape San Lucas, Mexico.
Remarks
Caprella californica [sensu lato] is one of the oldest species among the genus Caprella, originally reported from California in the 19th Century by Stimpson (1857). Mayer (1890) synonymized C. californica [sensu lato] with C. scaura f. δ from Hong Kong, China. However, in Mayer (1903), C. californica [sensu lato] was treated as C. scaura f. ξ californica, one of the 6 formae for C. scaura. Dougherty and Steinberg (1953) moved C. californica from subspecies to species level. McCain and Steinberg (1970) synonymized C. scaura scauroides from the western North Pacific and C. scaura spinirostris from Chile with C. californica [sensu lato] without legitimate reasons. Laubitz (1970) has provided the only sufficient description and illustrations for C. californica [sensu lato] from California.
The present description and illustrations for C. californica [sensu stricto] agree closely with those of Laubitz (1970) in: (1) the head of male possesses the anteriorly curved dorsal projection, (2) in the male, pereonite 5 is longer than pereonites 3 and 4, and (3) in the male, pereopod 7 is slender, and the merus is subequal in length with the propodus. As written in the remarks on C. scauroides, the difference between C. californica [sensu lato] from California and C. californica [sensu lato] from the Uwa Sea should be attributed to differences at the species level. Thus, C. californica [sensu lato] from California is described here as C. californica [sensu stricto] and C. californica [sensu lato] from the Uwa Sea is described as C. scauroides, a new rank of species (see remarks on C. scauroides).
Caprella californica [sensu stricto] is one of the most studied species of Caprellidea along the Pacific coast of the USA (see Saunders 1966; Keith 1969, 1971; Caine 1977; Page et al. 2006; Sirota and Hovel 2006; Carr et al. 2011). Carr et al. (2011) studied the epifaunal communities in eelgrass (Zostera marina Linnaeus, 1753) beds in San Francisco Bay, California. Of the five stations they studied, C. californica was the dominant species of Caprella spp. at Richardson Bay, near the entrance channel of San Francisco Bay. Caine (1977) categorized the feeding modes of the Caprellidea into 5 categories, that is, browsing, filter-feeding, predation, scavenging, and scraping. Feeding in C. californica is dominated by filter-feeding and scraping. Caprella californica prefers the bryozoan Bugula neritina (Linnaeus, 1758) over the red alga Polysiphonia pacifica Hollenberg, 1942 and a green alga (genus Viva), while C. equilibra Say, 1818 showed no preference between B. neritina and P. pacifica (Keith 1971). The preference of C. californica for B. neritina is based on its cryptic adaptations to that substrate.
-
Caprella scauroides Mayer, 1903 (Figs. 5 , 6 , 7 , 8 ) (Japanese name: Togewarekara-modoki)
Fig. 6 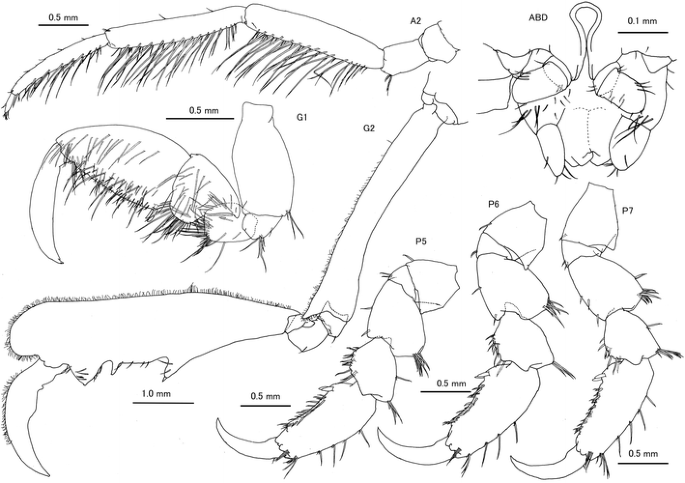
Caprella scauroides Mayer, 1903 from the Uwa Sea, southwestern Japan. Male “c”, AM-P.89082, 18.36 mm, A2, G1, G2, P5, P6 and P7; male “g”, AM-P.89084, 16.44 mm, ABD
Fig. 7 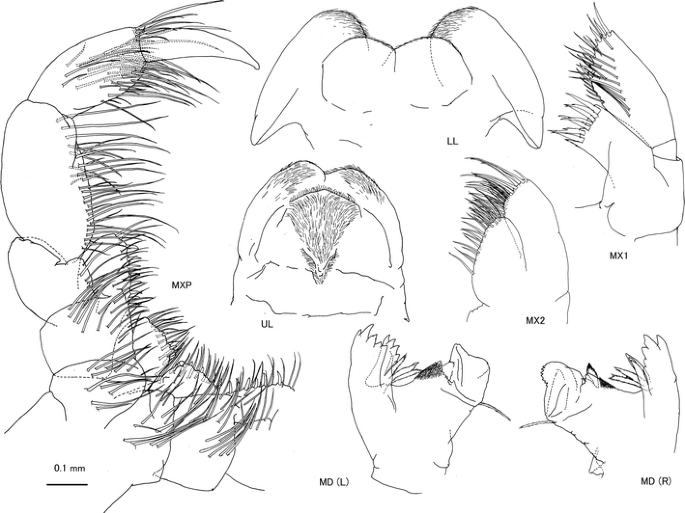
Caprella scauroides Mayer, 1903 from the Uwa Sea, southwestern Japan. Male “c”, AM-P.89082, 18.36 mm
Fig. 8 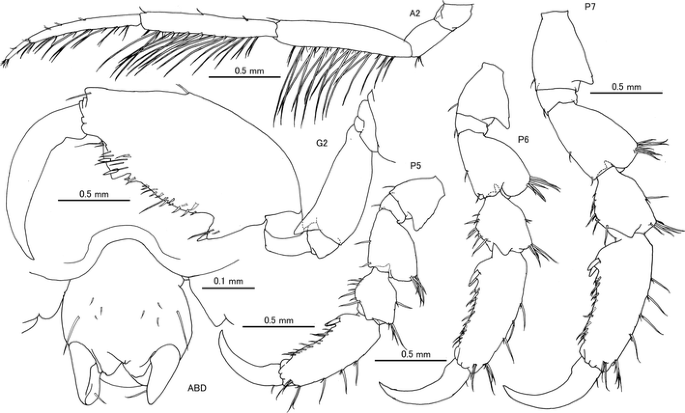
Caprella scauroides Mayer, 1903 from the Uwa Sea, southwestern Japan. Female “d”, AM-P.89083, 9.49 mm, A2, G1, G2, P5, P6 and P7; female “h”, AM-P.89085, 10.32 mm, ABD
-
Caprella californica: McCain and Steinberg 1970, 14 (in part); Arimoto and Kikuchi 1977, 91–92, fig 1b; Takeuchi 1995, 199, fig 21–181; Takeuchi 1999, 7; Aoki and Takeda 2006, 67; Lee and Hong 2009, 318; Montelli 2010, 726, fig 1.
-
Caprella (Spinicephara) californica: Arimoto 1976, 139–146, figs 75–77; Arimoto 1978, 27.
-
Caprella scaura f. δ (?): Mayer 1890, 72, pl 4 figs 43–44.
-
Caprella scaura f. scauroides Mayer, 1903: 118–119, pl 5 figs 16–18; Utinomi 1947, 77.
-
Caprella scaura scauroides: Krapp et al. 2006, 3.



Material examined
Male “c”, AM-P.89082, 33°11′N, 132°28′E, off Yusu, Uwajima in the Uwa Sea, Japan, pearl oyster aquaculture facility, 16 May 2006, coll. Takeuchi I; mature female “d”, AM-P.89083, 33°11′N 132°28′E, off Yusu, Uwajima in the Uwa Sea, Japan, pearl oyster aquaculture facility, 16 May 2006, coll. Takeuchi I; male “g”, AM-P.89084, 33°11′N 132°28′E, off Yusu, Uwajima in the Uwa Sea, Japan, pearl oyster aquaculture facility, 16 May 2006, coll. Takeuchi I; mature female “h”, AM-P.89085, 33°11′N 132°28′E, off Yusu, Uwajima in the Uwa Sea, Japan, pearl oyster aquaculture facility, 16 May 2006, coll. Takeuchi I; ca. 15 individuals, AM-P.89086, 33°11′N 132°28′E, off Yusu, Uwajima in the Uwa Sea, Japan, pearl oyster aquaculture facility, 16 May 2006, coll. Takeuchi I; 1 male, MCZ-126757, 33°11′N 132°28′E, off Yusu, Uwajima in the Uwa Sea, Japan, pearl oyster aquaculture facility, 16 May 2006, coll. Takeuchi I; 1 mature female, MCZ-126758, 33°11′N 132°28′E, off Yusu, Uwajima in the Uwa Sea, Japan, pearl oyster aquaculture facility, 16 May 2006, coll. Takeuchi I; ca. 20 individuals, MCZ-126759, 33°11′N 132°28′E, off Yusu, Uwajima in the Uwa Sea, Japan, pearl oyster aquaculture facility, 16 May 2006, coll. Takeuchi I.
Description
Male “c”, AM-P.89082. Body robust, 18.36 mm in body length. Pereonite 2 longest followed by pereonite 1. Head, 0.98 mm and pereonite 1, 3.49 mm; head and pereonite 1 partially fused; head with a forward-pointing dorsal projection; eye distinctive. Pereonite 2, 4.70 mm with a small mid-dorsal projection from 1/5 of posterior end, and a small ventral projection between gnathopod 2; height 1/3 × length. Pereonite 3, 2.48 mm with a small mid-dorsal projection and a dorsodistal projection and anterolateral projection. Pereonite 4, 2.19 mm with a small mid-dorsal projection and anterolateral projection. Pereonite 5, 2.59 mm with a mid-dorsal projection. Pereonite 6, 0.96 mm with 1 and a pair of mid-dorsal projections. Pereonite 7, 0.96 mm with a pair of mid-dorsal projections and a pair of dorsodistal projections.
Antenna
1, 0.65 × body length; peduncular article 1 robust; article 2 longest, 1.7 × article 1; article 3, 1.1 × article 1; flagellum with 17 articles, proximal article composed of 4 articles. Antenna 2, 0.4 × antenna 1; peduncular article 2 to flagellum article 1 with swimming setae.
Upper lip notched, wider than long, forming a pair of rounded projections, and finely setose. Lower lip well developed. Mandible left incisor with 5 teeth followed by lacina mobilis with 5 teeth and 3 accessory setal rows; molar well developed with a large flake-like projection and a long seta. Mandible right incisor with 5 teeth followed by lacina mobilis with 3 teeth and 2 accessory setal rows; molar well developed with a small molar flake and a long seta. Maxilla 1 outer plate with 7 stout apical setal-teeth; palp biarticulate; article 2, 5 × article 1 with a row of slender to robust marginal setae and 2 rows of lateral setae. Maxilla 2 inner plate oval with 2 lows of apical setae; outer plate with 2 rows of apical setae. Maxilliped lateral to basal part setose; basal endite (inner plate) round with 2 stout teeth and a row of apical setae; ischial endite (outer plate) subequal to inner plate with a rows of stout apical setae; palp 4 articulate; article 2 longest, setose on inner margin; article 3, 0.8 × article 4, setose on lateral to apical margin; palp article 4 (dactylus) falcate.
Pereon
Gnathopod 1; basis subequal with propodus with 3 posterodistal setae; merus to carpus densely setose; propodus subtriangular, 2 × width and densely setose on palm and lateral margin; palm begins near posterior margin with a pair of strong proximal spines; dactylus slightly curved distally.
Gnathopod 2 begins 1/5 along posterior margin of pereonite 2; basis subequal with pereonite 2, with dense fine setae and an anterodistal triangular projection; propodus subequal to basis and 3 × width; dorsal surface with dense fine setae and a large round apical projection; palm scarcely setose; proximal projection with a robust seta from 1/2 of proximal end; mid-palmar projection followed by a sinus and a large triangular projection; dactylus falcate, finely setose on dorsal margin.
Gill 3 oval 1.5 × width and subequal to length of pereonite 3. Gill 4 similar to gill 3.
Pereopods 5 to 7 stout; pereopod 5, basis with a distal triangular projection; merus 1.2 × basis and 0.6 × propodus, laterally expanded with several setae; carpus 0.8 × merus; propodus 2 × carpus with a pair of stout setae and ca. 10 pair of setae on palm; dactylus falcate. Pereopod 6 slightly longer, 1.1 × pereopod 5. Pereopod 7 subequal to pereopod 6; merus 1.5 × basis and 0.7 × propodus.
Pleon [Based on male “g”, AM-P.89084, 16.44 mm]
Uropod 1 uniramus with 7 lateral setae. Uropod 2 uniramus with an apical seta.
Female “d”, AM-P.89083.
Body length, 9.49 mm. Pereonite 2 longest followed by pereonite 3. Head length, 0.85 mm and pereonite 1, 0.62 mm; head with a forward-pointing projection; eye distinctive; pereonite 1 with a small dorsodistal projection; pereonite 2, 1.82 mm with a small mid-dorsal projection, a small dorsodistal projection and a ventral projection between gnathopod 2; pereonite 3, 1.69 mm with a mid-dorsal projection and anterolateral projection; pereonite 4, 1.43 mm with a mid-dorsal projection and dorsodistal projection; pereonite 5, 1.59 mm, with a mid-dorsal projection; pereonite 6, 0.78 mm with 1 and a pair of mid-dorsal projections; pereonite 7, 0.70 mm with a pair of anterodorsal projections. Antenna 1 0.6 × body length; peduncular article 2 longest; flagellum with 17 articles, proximal article composed of 2 articles. Antenna 2 slender, 0.6 × antenna 1.
Pereon
Gnathopod 2 begins 2/5 along anterior margin of pereonite 2; basis 0.5 × pereonite 2 with an anterodistal triangular projection; carpus triangular; propodus 1.5 × basis and 2.5 × width; dorsal surface convex with 2 setae with a small apical triangular projection; palm scarcely setose; proximal projection with 1 robust seta from 1/4 of proximal end; small mid-palmar projection followed by a small sinus and a triangular projection; dactylus falcate.
Pleon
[Based on female “h”, AM-P.89085, 10.32 mm]. Provided with 1 long seta followed by 3 small setae instead of uropod 1. Uropod 2 with an apical seta.
Distribution
Type locality: Hong Kong, China.
Other records: Aomori Prefecture to Kyoto Pref. along the coast of the Sea of Honshu Island, Seto Inland Sea, Fukuoka Pref. to Kumamoto Pref. of western coast of Kyushu Island, and Chiba Pref. to Miyazaki Pref. along the Pacific coast of Honshu, Shikoku and Kyushu Islands, Japan. South Jeolla Province (Jeollanam-do), Korea. Yellow Sea. The Port of Cairns, Queensland, Sydney Harbour, New South Wales, Hobsons Bay, Victoria, and Cockburn Sound, Western Australia, Australia.
Remarks
Mayer (1890) briefly described an unnamed forma (“forma δ (?)” in the caption of Plate 4) differing from C. scaura f. typica and C. scaura f. diceros, based on specimens collected from Hong Kong, China. Later, Mayer (1903) named this forma Caprella scaura f. scauroides with the description and the lateral view figures of 2 males and 1 mature female. The characteristics of male C. californica [sensu lato] from the Uwa Sea agree well with those of the larger male C. scaura f. scauroides in Mayer (1903; Plate 5 Fig. 18) in (1) the body somites are stout; (2) head possesses the straight, forward-pointing dorsal projection; (3) the propodus of gnathopod 2 possesses a round apical projection; (4) gills are oval, and (5) pereonite 5 is almost the same length as pereonites 3 and 4. The characteristics of mature females C. californica [sensu lato] from the Uwa Sea also agree favorably with C. scaura f. scauroides as illustrated in Mayer (1903; Plate 5 Fig. 16).
The present study indicated that C. californica [sensu lato] from the Uwa Sea, Japan, differs from C. californica [sensu lato] from California in the following characteristics:
-
1.
The body somites of C californica [sensu lato] from the Uwa Sea are more robust than C californica [sensu lato] from California in both sexes. The height of pereonite 2 in male C. californica [sensu lato] from the Uwa Sea is 1/3, while it is 1/4 in C. californica [sensu lato] from California.
-
2.
Male C. californica [sensu lato] from the Uwa Sea possess a straight, forward-pointing dorsal projection, while those from California possess an anteriorly curved dorsal projection.
-
3.
In male C. californica [sensu lato] from the Uwa Sea, the propodus of gnathopod 2 possesses a round apical projection, while the California specimen has a rectangular apical projection.
-
4.
In male C. californica [sensu lato] from the Uwa Sea, pereonite 5 is the same length as pereonites 3 and 4, while in the California specimen, pereonite 5 is longer than pereonites 3 and 4. In females, pereonite 2 is the longest among the body somites of C. californica [sensu lato] from the Uwa Sea, while in the California form pereonite 5 is the longest.
-
5.
In male C. californica [sensu lato] from the Uwa Sea, pereonite 7 has a pair of mid-dorsal projections, while in the California specimen, pereonite 7 has no mid-dorsal projections.
-
6.
In the Uwa Sea form, pereopods 6 and 7 are robust and slightly longer compared with pereopod 5; pereopod 7 is 1.3 times longer than pereopod 5 in males. The merus of pereopod 7 is shorter than the propodus. Conversely, in the California specimen, pereopods 6 and 7 are slender and much longer than pereopod 5; pereopod 7 is 2 times longer than pereopod 5 in males. The merus of pereopod 7 is the same length as the propodus.
Takeuchi (1989) described the body somites and appendages of each instar of 4 species of Caprella reared in the laboratory. He revealed that the characteristics of antennae 1, 2, and pereopods 5–7 are rather stable in both sexes and growth stages, compared with other characteristics including the ratio of each pereonite length and gnathopod length to the body length. The difference between C. californica [sensu lato] from the Uwa Sea and C. californica [sensu lato] from California should be attributed to differences at the species level. Thus, C. californica [sensu lato] from California is described as C. californica [sensu stricto] (see remarks on C. californica) and C. californica [sensu lato] from the Uwa Sea was described as C. scauroides Mayer, 1903, a new rank of species.
Caprella californica [sensu lato] was one of the dominant species of the genus Caprella along the coast of Japan. Arimoto (1976) reported that C. californica [sensu lato] distributed from the western part of Kyushu to the Tsugaru Strait along the coasts of the Sea of Japan, Seto Inland Sea coasts and the southeast part of Kyushu to Tateyama Bay along the Pacific coasts of Japan, and the middle of the Yellow Sea. C. californica [sensu lato] was listed as one of the dominant species of Caprella spp. from Amakusa, western Kyushu (Aoki 1999) and as one of the dominant epifaunal species on the gorgonian Melithaea flabellifera (Kükenthal, 1908) at the Izu Peninsula, central Japan (Kumagai and Aoki 2003). In 2001, the usage of Caprella spp. to monitor temporal and spatial changes of butyltin concentrations in shallow water ecosystems was proposed (Takeuchi et al. 2001). The butyltin concentrations in seawater and Caprella spp. from the Uwa Sea and Uranouchi Bay, where the dominant industry is aquaculture, was equal to or significantly higher than samples from the western part of the Seto Inland Sea, a major, heavy-industrial area in western Japan (Murai et al. 2005). Caprella californica [sensu lato] was one of the dominant species of Caprella spp. in this survey. Matsuo et al. (2010) also reported the occurrence of C. californica [sensu lato] as the dominant species of Caprella spp. inhabiting fish and pearl oyster aquaculture facilities along the coasts of the Uwa Sea during a survey on nitrogen loading from fish aquaculture facilities. Caprella californica [sensu lato] in the above studies should henceforth be regarded as C. scauroides.
Caprella californica [sensu lato] was recently reported from the Australian coast (Montelli 2010). C. californica [sensu lato] was first recorded from Sydney Harbour, New South Wales in 2002, and Cockburn Sound, Western Australia, Hobsons Bay, Victoria, and the Port of Cairns, Queensland. Based on the lateral view photograph of a male (Montelli 2010; p 726), the Australian C. californica [sensu lato] is more similar to C. scauroides from the Uwa Sea, Japan, than to C. californica [sensu stricto] from California in robust body somites, the straight, forward-pointing dorsal projection on the head, round apical projection on the propodus of gnathopod 2, and the similar lengths of pereonites 3, 4 and 5.
Mayer (1890) briefly described C. scaura f. spinirostris, supplying figures of the head and propodus of ganthopod 2. Mayer (1903) reported the presence of a projection in the ventral face of pereonite 2 in small males and paired projections on pereonite 5 on old males of C. scaura f. spinirostris. Krapp et al. (2006) also mentioned that C. scaura spinirostris has a ventral spine, unlike C. scaura diceros, C. scaura cornuta and C. scaura typica. As written in the Introduction, McCain and Steinberg (1970) synonymized C. scaura f. spinirostris with C. californica [sensu lato]. Guerra-García and Thiel (2001) reported “Caprella scaura” from Coquimbo, northern-central Chile, which is the type locality of C. scaura f. spinirostris. They simply described that “C. scaura” from Coquimbo possesses the ventral projection between gnathopod 2 on pereonite 2. While the characteristics of gnathopod 2 and pereonite 5 are unclear, their description indicates that “C. scaura” from Coquimbo might actually be C. scaura f. spinirostris. Nevertheless, in the synonym list for “C. scaura” in Guerra-García and Thiel (2001) “C. californica” and C. scaura f. spinirostris was not included. Thus, further detailed studies with descriptions and figures are necessary before the taxonomic status of “Caprella scaura” from Coquimbo, Chile can be clarified.
References
Aoki M (1999) Morphological characteristics of young, maternal care behaviour and microhabitat use by caprellid amphipods. J Mar Biol Assoc U.K. 79:629–638
Aoki M, Takeda M (2006) Caprellid amphipods from the coastal waters of Shimoda, Izu Peninsula, Central Japan. Mem Natl Sci Mus Tokyo 41:65–70
Arimoto I (1976) Taxonomic studies of caprellids (Crustacea, Amphipoda, Caprellidae) found in the Japanese and adjacent waters. Spec Publ Seto Mar Biol Lab 3:i–v+1–229
Arimoto I (1978) Caprellids (Amphipoda, Caprellidea) from Kushimoto (Honshu) in Japan. Proc Jpn Soc Syst Zool 14:25–28
Arimoto I, Kikuchi T (1977) Caprellids obtained in the vicinity of the Amakusa Islands, west Kyushu. Publ Amakusa Mar Biol Lab Kyushu Univ 4:91–98
Boeck A (1872) B idrag til Californiens Amphipodefauna. Forhandlinger I videnskabs selskabet I Christiania: 33–51
Caine EA (1977) Feeding mechanisms and possible resource partitioning of Caprellidae (Crustacea: Amphipoda) from Puget Sound, USA. Mar Biol 42:331–336
Carr LA, Boyer KE, Brooks JA (2011) Spatial patterns of epifaunal communities in San Francisco Bay eelgrass (Zostera marina) beds. Mar Ecol 32:88–103
Dougherty C, Steinberg JE (1953) Notes on the skeleton shrimps (Crustacea: Caprellidae) of California. Proc Biol Soc Wash 66:39–50
Guerra-García JM, Thiel M (2001) The caprellid fauna (Crustacea: Amphipoda: Caprellidea) from the coast of Coquimbo, Northern-central Chile with a taxonomic key for species identification. Rev Chil Hist Nat 74:873–883
Keith DE (1969) Aspects of feeding in Caprella californica Stimpson and Caprella equilibra Say (Amphipoda). Crustaceana. 16:116–124
Keith DE (1971) Substrate selection in caprellid amphipods of Southern California, with emphasis on Caprella californica Stimpson and Caprella equilibra Say (Amphipoda). Pac Sci 25:387–394
Krapp T, Lang C, Libertini A, Melzer RR (2006) Caprella scaura Templeton, 1836 sensu lato (Amphipoda: Caprellidae) in the Mediterranean. Org Divers Evol 3(6 Electr Suppl):1–18
Kumagai NH, Aoki M (2003) Seasonal changes in the epifaunal community on the shallow-water gorgonian Melithaea flabellifera. J Mar Biol Assoc U.K. 83:1221–1222
Laubitz DR (1970) Studies on the caprellidae (Crustacea, Amphipoda) of the American North Pacific. Publ Biol Oceanogr 1:v–vii+1–89
Lee K-S, Hong S–S (2009) Caprellid fauna (Amphipoda: Caprellidae) of Goheung Peninsula, Korea. Korean J Syst Zool 25:317–323
Martin DM (1977) A survey of the family Caprellidae (Crustacea, Amphipoda) from selected sites along the northern California coast. Bull South Calif Acad Sci 76:146–167
Matsuo H, Matsuka N, Omori K, Koizumi Y, Takeuchi I (2010) Difference in the stable nitrogen isotope ratio of Sargassum piluliferum (Phaeophyceae: Fucales) associated with fish and pearl oyster aquaculture facilities. Fish Sci 76:147–153
Mayer P (1882) Monographie: Caprelliden. Fauna und flora des golfes von Neapel und der angrerzenden Meeres-Abschnitte 6:i–x, 1–201, pls I–X
Mayer P (1890) Monographic: nachtrag zur den caprelliden. fauna und flora des golfes von neapel und der angrerzenden Meeres-Abschnitte 17:i–vii, 1–157, pls I–VII
Mayer P (1903) Die Caprelliden der Siboga-Expedition. Siboga-Expeditie 34:1–160, pls. 1–X
McCain JC, Steinberg JE (1970) Amphipoda 1 Caprellidea 1. Fam. Caprellidae. Gruner HE, Holthuis LB (ed.) Crust Catalog 2:1–78
Montelli L (2010) The recent geographical expansion of Caprella californica (Caprellidea: Caprellidae) around the coastline of Australia. Biol Invasions 12:725–728
Murai R, Takahashi S, Tanabe S, Takeuchi I (2005) Status of butyltin pollution along the coasts of Western Japan in 2001, 11 years after partial restrictions on the usage of tributyltin. Mar Pollut Bull 51:940–949
Page HM, Dugan JE, Culver CS, Hoesterey JC (2006) Exotic invertebrate species on offshore oil platforms. Mar Ecol Prog Ser 325:101–107
Saunders CG (1966) Dietary analysis of caprellids (Amphipoda). Crustaceana 10:314–316
Sirota L, Hovel KA (2006) Simulated eelgrass Zostera marina structural complexity: effects of shoot length, shoot density, and surface area on the epifaunal community of San Diego Bay, California, USA. Mar Ecol Prog Ser 326:115–131
Stimpson W (1857) The Crustacea and Echinodermata of the Pacific shores of North America. Boston J Nat Hist 6:444–532
Stimpson W (1873) On some Californian Crustacea. Proc Calif Acad Sci 1:95–99 Second Edition
Takeuchi I (1989) Taxonomic and ecological studies of the Caprellidea (Crustacea, Amphipoda) inhabiting the Sargassum zone. Dissertation, Graduate School of Agriculture, The University of Tokyo (in Japanese).
Takeuchi I (1995) Suborder Caprellidea. In: Nishimura S (ed) Guide to seashore animals of Japan with color picture and keys, vol 2. Hoikusha, Osaka, pp 193–205 (in Japanese)
Takeuchi I (1999) Checklist and bibliography of the Caprellidea (Crustacea: Amphipoda) from Japanese waters. Otsuchi Mar Sci 24:5–17
Takeuchi I, Takahashi S, Tanabe S, Miyazaki N (2001) Caprella watch; a new approach for monitoring butyltin residues in the ocean. Mar Environ Res 52:97–113
Utinomi H (1947) Caprellidea of Japan and adjacent waters. Seibutu Suppl 1:68–82
Watling L, Carlton JT (2007) Caprellidae. In: Carlton JT (ed) The Light and Smith manual intertidal invertebrates from central california to oregon. Fourth edition, completely revised and expanded. University of California Press, Berkeley and Los Angeles, pp 618–629
Acknowledgments
We thank Mr. A. Baldinger and Ms. C. Piotrowski for loaning the specimens of Caprella californica [sensu stricto], collected from California, USA, Mr. K. Furuya and Dr. S. Suzuki for support in collecting C. scauroides from the Uwa Sea, Japan, and Ms. J.H.C. Lim for her critical reading and constructive comments on the MS.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by H.-D. Franke.
Rights and permissions
About this article
Cite this article
Takeuchi, I., Oyamada, A. Description of two species of Caprella (Crustacea: Amphipoda: Caprellidae) from the North Pacific; C. californica Stimpson, 1857 and C. scauroides Mayer, 1903, with a new appraisal of species ranking for C. scauroides . Helgol Mar Res 67, 371–381 (2013). https://doi.org/10.1007/s10152-012-0329-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10152-012-0329-9