- Original Article
- Published:
Leucothoid and Maerid Amphipods (Crustacea) from deep regions of the North Atlantic
Helgoland Marine Research volume 67, pages 383–396 (2013)
Abstract
From amphipod collections from the deep Norwegian Sea, three species are here redescribed: Maera loveni (Bruzelius 1859) (Maeridae), Leucothoe aff. spinicarpa (Abildgaard, 1789) and L. uschakovi Gurjanova, 1951 (Leucothoidae). A key to all Atlantic and Mediterranean Leucothoe species (males only) is provided.
Introduction
The amphipod fauna of the deep Norwegian Sea is as yet very insufficiently known. Most of our knowledge still rests on the data from the Norske Nordhavs Expedition (Sars 1885) and the Danish Ingolf Expedition (Stephensen 1923, 1925, 1931, 1944). In recent years, much marine material has been collected during the BioFar and BioIce programmes, but the amphipod collections from these programmes have not yet been worked out. In addition, Norwegian marine biologists, primarily Torleiv Brattegard, the late Torleif Holthe and Jon Arne Sneli, have during the 1970s and 1980s brought together large collections of marine animals from the continental slope and abyssal plain of the Norwegian Sea; this material rests in the collections of the Zoological Museum, Bergen.
In the year 2009, Vader and Tandberg, with financial support from the Norwegian Academy of Sciences, organized an International Amphipod Workshop in Skibotn, N. Norway, where part of these collections, that is material from deeper than 1,000 m, was sorted and in many cases preliminarily identified. In 2011, the present authors, again with support from the Norwegian Academy of Sciences, got the opportunity to describe the Maeridae and Leucothoidae from this material in more detail, and we present here the results of this study.
Material and methods
Samples were taken mainly by RP bottom sledge. The amphipods were fixed using 3–5 % formalin, then washed, preserved in 70 % alcohol and sorted under a dissecting microscope. Slides were prepared with glycerine and/or Faure’s medium. Pencil drawings were done using various compound microscopes with camera lucida; the transfer or “inking” was done by hand as well as using the program Adobe Illustrator.
All examined material is deposited at the Crustacean collection of the Zoological Museum, Bergen.
Systematics
Family Maeridae Krapp-Schickel, 2008
-
Maera loveni (Bruzelius, 1859) (Figs. 1, 2)
Fig. 1 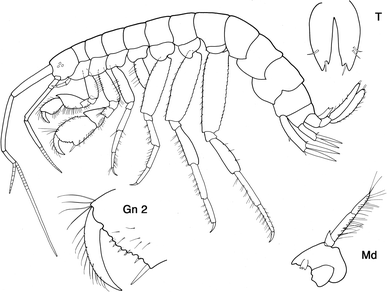
Maera loveni (Bruzelius, 1859): habitus; Gn 2 gnathopod 2; Md mandible, T telson
Fig. 2 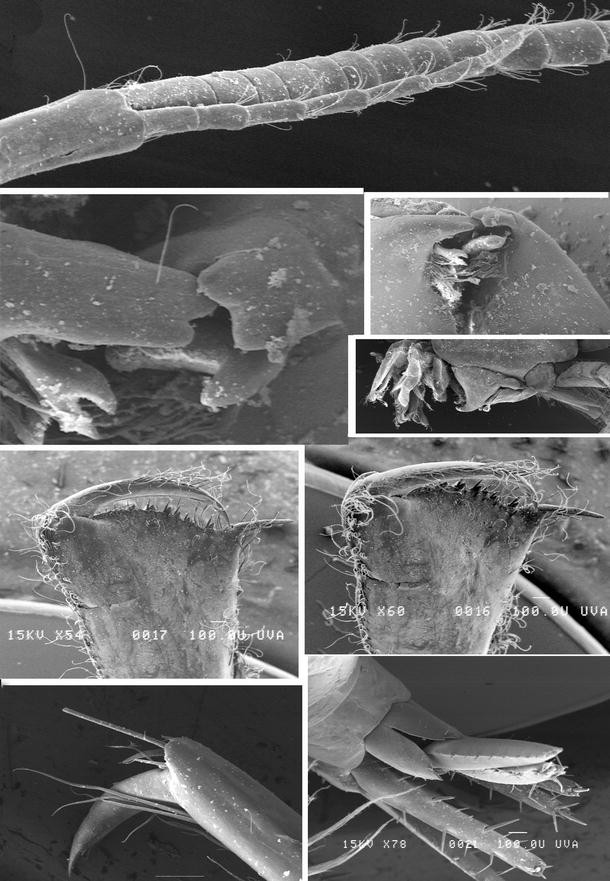
Maera loveni (Bruzelius, 1859): SEM pictures. Upper half first antenna with accessory flagellum. Directly underneath: mouthpart-details (on left and right side the two mandibles working together, on right side an overview of the mouthpart bundle, laterally seen). Lower half: second gnathopod in two different positions. Lowest left: dactylus and propodus of peraeopod 7. Lowest right: telson and uropods seen from dorsally
-
Gammarus loveni Bruzelius 1859: 59
-
Maera loveni Bate 1862: 193; Norman 1869: 416; Sars 1895: 519, pl. 182; Stebbing 1906: 438; Shoemaker 1930: 116; Schellenberg 1942: 44; Gurjanova 1951: 757–758, fig. 526; Lincoln 1979: 286, fig. 133; Krapp-Schickel and Jarrett, 2000: 34, fig. 4.
-
Maera tenera G.O. Sars, 1885 (cf. also Stephensen 1940: 311 who already proposed this synonymy)
-
Maera prionochira von der Brüggen 1907: 230, Figs. 5, 6, 7 (figure captions are most probably switched: what is labelled as Gn 1 is the normal Gn 2, and what is labelled as Gn 2 must be an anomaly. But Ep 3 and T shape are matching).


Type locality
Bohuslän, Koster Islands, Gullmarfjord, “locis profundioribus”.
Material examined
1 male 34 mm, 83.06.03.2, 1.220 m depth, 60°12,1′N, 06° 37,5′W 1983 3/VI.
Diagnosis
Largest member of the Maera sensu stricto—group; diagnostic differences are the very little visible ommatidia of the small eyes, long flagellum in A 1, the anteriorly acutely lengthened Cx 1, the long spine on the palmar corner of Gn 2, the posterodistal corner of Ep 1–3 with upturned acute tooth, and the asymmetrically incised lobes in the telson with one spine inserted on the inside.
Redescription (using the original one by Bruzelius, completed by the allometric differences of the present specimen):
Body
Uncommonly elongate and narrow, back completely smooth. [Length of type material 20 mm, up to 37 mm after Shoemaker 1930: 334; in the present specimen 34 mm].
Head
Lateral cephalic lobe regularly rounded, anterodistal corner acute. Eyes small [with scattered ommatidia, which are pale and scarcely visible in alcohol].
A1 reaching segment 4 of the abdomen, about twice as long as A 2, regularly beset with short setae; peduncle art 1 long and narrow, but thicker and shorter than art 2; [art 3 about 1/3 length of art 2]; flagellum with about 30 arts, acc. flag. with 7–8 arts.
A 2 ped art 3 much shorter and somewhat thicker than art 4, art 4 subequal art 5 [here art 4 > art 5], art 4 longer than end of A 1 peduncle art 1; art 5 does not reach the end of A1 ped art 2 [does reach it here]; flagellum somewhat shorter than peduncle art 5 [here subequal], consists of 10–12 arts [here 13].
Mouthparts
Mandibles strong, with high molar; palp with 3 arts, the second the longest, art 1 the shortest and with tooth-shaped prolongation. Mx1 IP narrow, distally with 3–4 plumose setae; [OP with bifurcate robust spines]; Mx 2 lobes elongate, densely beset with long marginal setae [partly plumose; on IP also facial setae]. Mxp IP reaches about half of palp art 2, distally with dense setae; OP (=ischium) reaches about 1/3 of palp art 2 (=carpus), along inner margin strong spines which are toothed on one side, distally with setae; art 2 of the palp is the longest and is also densely beset with spines and setae. UL rounded, LL divided into large rounded lobes with short processes anterodistally.
Peraeon
Coxal plates short, Cx 1, 2 smaller than Cx 3, 4 [here clearly the opposite]. Cx 1 anterodistal corner acutely produced, all other coxae rounded.
Gn 1 much smaller than Gn 2
Carpus subequal to propodus [here longer], anterodistal corner acute, posterior margin rounded; anterior margin almost without setae, a number of oblique rows of densely long setae along the posterior margin; [propodus distally widening, posterior margin regularly rounded]; also facial setae on both sides. Dactylus strong, outer margin with long and fine setae, but no spines.
Gn 2 carpus [triangular]
Much smaller than propodus, on posterior margin very dense setae; propodus almost rectangular, proximal part somewhat narrower than distal one. Palm slightly convex, with 7–8 small acute humps beset with shorter spines and longer setae; palmar corner with a strong, straight and acute spine, posterior margin with dense short setae.
P 3 somewhat longer than P 4
Subsimilar: basis long and narrow, ischium short, merus, carpus and propodus gradually shorter; dactylus short [robust].
P 5–7 uncommonly narrow bases (in the type material of 20 mm)
Distally narrowing [in our specimen of 34 mm bases not so narrow], [anterior margin beset with short setae, posterior margin finely serrated; in P 5 basis proximally rounded, in P 6, 7 basis with posteroproximal lobe]; dactyli strong, convex and acute.
Pleon
Ep 1, 2, 3 all with small upturned sharp tooth on posterodistal corner.
U 1 peduncle longer than subequal rami and reaching somewhat further than U 2, with one strong spine along half of anterior margin and two spines distally; rami beset with short spines along the margins and distally; U 2 peduncle shorter than subequal rami, U 3 reaching much further than U 1, 2, rami flattened, subequal in length, margins with shorter, distal end with longer setae and fine spines.
Telson deeply cleft, but not quite to the basis, lobes diverging, distally narrowing and with short V-shaped incision [inner arm of this “V” much shorter], beset with one strong spine [which is subequal or somewhat longer than outer arm of incision].
No sexual differences observed; the female has only slightly shorter appendages.
Colour
Pale.
Distribution
North Atlantic, American and European coasts; Arctic Ocean; Greenland; Iceland; European coasts from northern Norway to British Isles. Depth range as yet only known from 6 to 400 m. Now down to 1,220 m.
For a key to all 12 Maera species worldwide (M. anoculata, M. danae, M. edwardsi, M. fusca, M. grossimana, M. hirondellei, M. loveni, M. pachytelson, M. schieckei, M. similis, M. sodalis, M. tinkerensis) see Krapp-Schickel and Jarrett 2000: 29.
Leucothoidae Dana, 1852
-
Leucothoe aff. spinicarpa (Abildgaard, 1789) (Figs. 3, 4)
Fig. 3 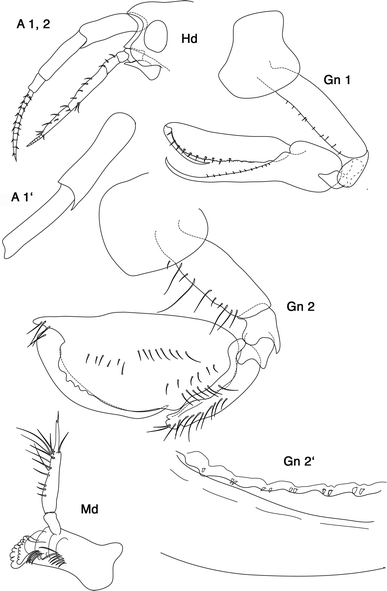
Leucothoe aff. spinicarpa (Abildgaard, 1789): A 1, 2 antenna 1, 2; A 1′ first antenna peduncle article 1, 2 enlarged. Hd head. Gn 1, 2, Gn 2′ parts of dactylus and propodus enlarged. Md mandible
Fig. 4 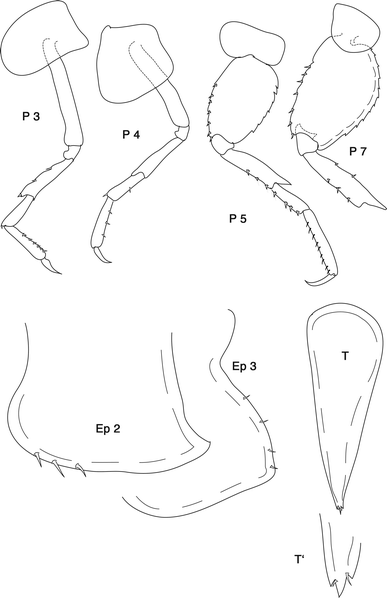
Leucothoe aff. spinicarpa (Abildgaard, 1789): P 3–7 peraeopods. Ep 2, 3 = epimeral plates 2, 3. T telson, T′ distal end of telson enlarged
-
Krapp-Schickel 1989; Krapp-Schickel and Menioui 2005, 64–66; Crowe 2006: 57–68.


Material examined
1 ? fem. 6 mm, 4 immatures 4–5 mm, 1,542–1,560 m depth, 62°28.1′N, 14° 13.4′W, Stn 83.06.06.1, 1983 6/VI.
Diagnosis
Eyes oval. Mandibular palp long and narrow, art 3 half length of art 2. Cx 1 inferior margin smooth, about as long as wide. Gn 1 carpus distal part about 6× longer than wide, dactylus reaching about 0,3× of propodus length. P 3, 4 with very narrow basis, P 5–7 basis ovally broadened, with regularly rounded hind margin, ratio length to width 2.5: 4 or 3.5: 5. Ep 2 posterodistally with upturned tip, Ep 3 distoposterior corner with right angle, blunt, anterior corner lacking setae.
Description
Length 4–6 mm.
Head
Anterior margin rounded, anterodistal margin rectangular with rounded corner. Mid-cephalic keel with acute projection. Rostrum small.
Eyes oval
Antenna 1 one-third of body length, flagellum 11- articulate, peduncle art 1 width proximally less than twice article 2, disto-inferiorly with acute tooth, length art 1 subequal art 2, art 3 about 1/3 of art 2, acc. flagellum not seen.
Antenna 2 one-third of body length, subequal in length to antenna 1, peduncle art 4 > art 5, flagellum 6 arts.
Mouthparts
Mandibles lacking molars, palp 3–articulate, ratio of articles 1–3 is about 1: 2,8: 1.4, art 2 with 8–9 long lateral and 1 distal setae, art 3 with 1 distal seta, incisors strongly dentate.
Peraeon
Cx 1–4 relative widths 1.0: 1.2: 0.8: 1.
Cx 1 smooth, length and width subequal; anterior margin smooth, excavate, anterodistal margin produced, anterodistal corner about 120°, distal margin regularly rounded, smooth, posterior margin excavate, facial setae absent.
Gn 1 basis not inflated, with small tiny setae on anterior margin; ischium smooth; carpus distal part linear, length about 6× width; propodus straight, palm dentate with 9 short spines; dactylus smooth, reaching about 0.3× propodus length.
Cx 2 about as long as wide, subquadrangular, much wider than Cx 3, distally smooth; anterior margin straight, anterodistal corner rectangular, inferior margin rounded, posterior margin straight, facial setae absent.
Gn 2 basis slightly inflated, on anterior margin 10 setae of different length; carpus nearly reaching half propodus length, curved, distally truncate, densely setose; propodus distally with tooth-shaped prolongation, palm convex with many low humps, with ?2 mediofacial setal rows, with a few submarginal setae, proximally near dactylus end widening with clear corner of about 120°; dactylus curved, both margins smooth, bare, reaching somewhat more than half of propodus length.
Cx 3 length > width, smooth, bare, anterior and posterior margin straight, distally rounded, facial setae absent.
Cx 4 wider than long, smooth, bare, anterior margin somewhat convex, distal margin scarcely rounded, posterior margin shorter than anterior one, excavate, facial setae absent.
P 3, 4 basis very narrow, about the width of merus and up to 6–7× the width of basis; dactylus reaching nearly half length of propodus, posterior margins with short and thin spines.
Cx 5–7 facial setae absent.
P 5–7 similar, bases length: width ratio about 1.4–1.6, anterior margins with slight serrations and small weak spines, posterior margins strongly serrate.
Pleon
Ep 1 posteroventral corner rounded. Ep 2 with spines on distal margin, posteroventral corner acutely produced. Ep 3 posteroventral corner blunt and rectangular, distally bare, but some short spines on posterior margin.
U 1–3 relative lengths 1.0: 0.8: 1.1.
Telson
Ratio length: width about 3, tip tridentate because of two indentations near distal end, with a short seta inserted in each one.
Remarks
These specimens undoubtedly belong to the “spinicarpa”-group.
We started out with the supposition that material of more than 1,500 m depth cannot be L. spinicarpa which primarily is a shallow water species. Sars (1891) writes “30–150 fathoms”, while Lincoln (1979) records the species as “occasionally intertidally, down to 600 m”. Our material contains no specimens with signs of sexual maturity, and we know extremely little about the life span, variability or allometry in L. spinicarpa. The species has been redescribed by Krapp and Menioui (Krapp-Schickel and Menioui 2005) and by Crowe (2006) on the basis of topotypical material; Crowe also designed a male neotype of 13 mm, deposited in the Zoological Museum of Oslo.
L. spinicarpa (Abildgaard, 1789) is one of the earliest amphipods to be formally described, and the description is correspondingly vague. Shortly after also the species L. articulosa Montagu, L. denticulata A. Costa and L. antarctica Pfeffer were described, all morphologically quite similar to L. spinicarpa and all for a long time synonymized with that species. L. denticulata was re-established and fully described by Krapp-Schickel and Menioui (2005), and the redescription of L. antarctica is in preparation, but a modern redescription of L. articulosa, also revived by Krapp-Schickel and Menioui (loc. cit.), is still lacking.
Sars (1890–1895) revised the species and described L. spinicarpa from Norway, synonymizing L. articulosa (type locality S-Devonshire, type material also lost) with it. But in his illustration on pl. 101 (sub male L. articulosa of 16 mm), Ep 3 has a small but well-defined posterodistal tooth-shaped tip, which is still more acutely illustrated in the drawing of L. articulosa by Bate and Westwood 1863 Fig. 3, while Lincoln 1979 describes Ep 3 in his material (sub L. spinicarpa) as “quadrate OR with minute blunt tooth”; in the illustration of the neotype, Ep 3 is posterodistally rectangular.
Crowe (2006) specified several characters useful for comparison within the “spinicarpa-group”: the number of flagellum articles on the antenna 1 and 2; the anterior setation of the basis and the mediofacial setae on the propodus of gnathopod 2; the propodus and dactylus length of pereopods 3–7; the shape and setation of epimera 3 and the telson.
Our material (4–6 mm) is not fully mature, and therefore a number of these characters cannot be applied here, for example the antennar articles. The deepwater specimens deviate in a few points from Crowe’s redescription of L. spinicarpa: Gn 2 carpus is distally rounded (as in females and juveniles of L. spinicarpa, but differing from adult males). Ep 1 lacks anterodistal setae in our material, and the mediofacial row (or 2 shorter rows?) of setae on the propodus of Gn 2 has a quite different shape than in Crowe‘s material.
As our material does not contain obvious males and none of the specimens seems to be fully adult, we refrain from describing a new species and design our material as L. aff. spinicarpa.
-
Leucothoe ushakovi Gurjanova, 1951 (Figs. 5, 6, 7)
Fig. 5 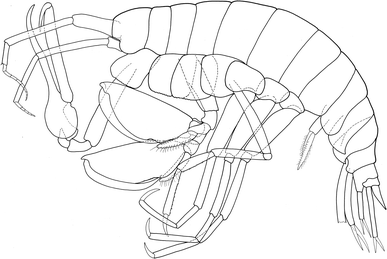
Leucothoe ushakovi Gurjanova 1951: habitus
Fig. 6 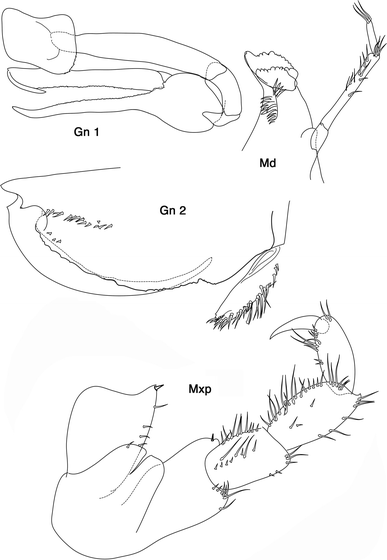
Leucothoe ushakovi Gurjanova 1951: Gn 1, 2 gnathopod 1, 2; Md mandible, Mxp maxilliped
Fig. 7 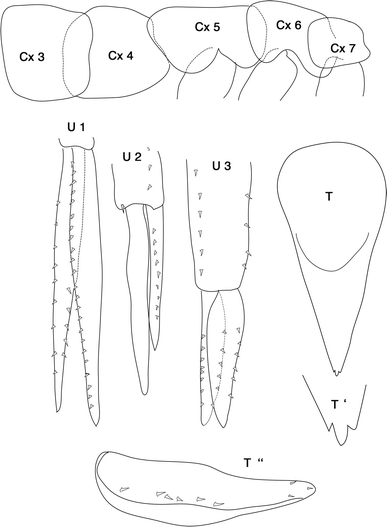
Leucothoe ushakovi Gurjanova 1951: Cx 3–7 = coxal plates; U 1–3 = uropods; T telson, T′ and T″ enlargement of telson distally and laterally
-
Gurjanova 1951: 487–488, fig. 320 A, B



Material examined
10 specimens of 8–27 mm length, 3,892 m depth, 65° 39,9′N, 04° 35,8′W, Stn 83.06.09.1, 1983 9/VI.
Type locality
Greenland Sea.
Diagnosis
Eyes apparently absent in fixed material. Mandibular palp long and narrow, art 3 up to 2.5× shorter than art 2. Cx 1 inferior margin weakly serrate, ratio length: width in large animals up to 2. Gn 1 carpus distal part about 10× longer than wide, dactylus reaching 0.25× of propodus length. P 3–7 with very narrow and similar basis, ratio length to width up to 4 in P 5–7, up to 7–8 in P 3, 4. Ep 2, 3 posterodistally with upturned tooth. U 1–3 peduncle always longer than rami.
Redescription
Length up to 34 mm.
Head
Anterior margin rounded, anterodistal margin rectangular with rounded corner. Ventral cephalic keel anteroventral margin without projection. Rostrum acute, small.
Eyes apparently absent in fixed material.
Antenna 1 one-third of body length, flag 6–15 articulate, ped art 1 width proximally about twice art 2, length art 1< art 2, art 3 half length art 2, acc. flag with 1 small art.
Antenna 2 one-third of body length, subequal in length with A 1, ped art 4> art 5, flag 6–10 arts.
Mouthparts
Mandibles lacking molars, palp 3–articulate, ratio of articles 1–3 is about 2: 6.5: 2.7 or 1.8: 6.2: 3, art 2 with 8–9 long lateral and 3 distal setae, art 3 with 2–3 distal setae, incisors strongly dentate; left mandible lacinia mobilis large, strongly toothed, with 16 raker spines; right mandible lacinia mobilis small, with 16 raker spines.
Lower lip inner lobes fused, bare; outer lobes with small gap.
Mx 1 palp with 2 arts, with 2 distal setae; outer plate with distal spines and setae, inner plate naked. Mx 2 inner plate with many thin marginal and a few facial setae; outer plate with few distal setae. Mxp inner plates fused, distal margin with a v-shaped indentation; outer plate triangular, reaching 0.3× palp art 1, with one subdistal spine; palp with 4 arts, art 2 and art 3 with marginal and facial setae, dactylus <art 3.
Peraeon
Cx 1–4 relative widths 1.0: 1.3: 1.3: 1.5.
Cx 1 smooth, in smaller specimens ratio length to width about 11: 8, in larger ones up to 2: 1; anterior margin smooth, anterodistal margin produced, rounded, distal margin straight, with some dentations, posterior margin excavate, facial setae absent.
Gn 1 basis scarcely inflated, naked; ischium with posterior setae; carpus distal part linear, length about 10× width, proximal margin finely serrated; propodus straight, palm dentate with 10 short setae; dactylus smooth, reaching 0.25× propodus length.
Cx 2 about as long as wide, subquadrangular, wider than Cx 3, distally weakly dentate; anterior margin straight, anterodistally rectangular, inferior margin straight, posterior margin straight, facial setae absent.
Gn 2 basis slightly inflated, naked; carpus 0.3× propodus length, curved, distally truncate, densely setose; propodus distally with tooth-shaped prolongation, posterior margin smooth, with 1 mediofacial setal row, with a few submarginal setae, palm convex and dentate, proximally near dactylus end widening with angle of about 120°; dactylus curved, outer margin smooth, bare, inner margin with rounded humps, reaching 0.75× propodus length.
Cx 3 length subequal width, anterodistal corner overriding distal face of Cx 2, smooth, bare, anterior margin straight, anterodistal corner rounded, facial setae absent.
Cx 4 wider than long, smooth, bare, anterior margin somewhat convex, distal margin evenly rounded, posterior margin excavate, facial setae absent.
P 3, 4 basis very narrow, about the width of merus and up to 7–8× the width of basis; dactylus reaching 1/3 of propodus, posterior margins with short and thin spines.
Cx 5–7 facial setae absent.
P 5–7 very similar, bases length: width ratio about 4, anterior margins with very slight serrations and small weak spines, posterior margins smooth.
Pleon
Ep 1 with 1 spine on inferior margin, Ep 2, 3 bare. Ep 1 posteroventral corner rounded, Ep 2, 3 posteroventral corner acutely produced.
U 1–3 relative lengths 1.0: 0.8: 1.1. U 1 peduncle nearly twice the outer ramus which is about 0.9× of inner ramus length. U 2 peduncle > inner ramus length, outer ramus length 0.8× inner ramus length. U 3 peduncle about twice inner ramus length, outer and inner ramus length subequal. U 1–3 margins of peduncle, inner and outer rami all with short spines.
Telson
Twice as long as wide, apex with very small V-shaped excavation in the acute tip, with submarginal short spines.
No sexual differences observed.
Colour
In alcohol somewhat yellowish, body semi-transparent.
Distribution
Greenland Sea at the depth of 3,000 m (2 specimens, types). Norwegian Sea at 65°N and 04° W, 3,892 m depth (presently studied material)
Remarks
Gurjanova (1951) reports about the type material that “Gn1 propodus is longer than basal article”, which is not the case in our material, obviously due to the propodus growing allometrically. About the telson, she described that the lateral margins are “completely smooth”, which was the case in some of the presently studied specimens, while others had some submarginal small spines.
Remarks on the genus Leucothoe
The amphipod genus Leucothoe is extremely easy to recognize as such, but its many species are differentiated from each other by only subtle characters, and this has traditionally led to a whole-sale lumping of many species under just a few names, esp. sub Leucothoe spinicarpa (Abildgaard, 1789). In the last years, it has become clear that the genus, with many of its species living commensally inside sponges, ascidians and even bivalve mollusks, is a very speciose one, and especially in the Pacific Ocean, there are still large numbers of as yet undescribed species (cf White and Reimer 2012a, b, c). The present identification key, which is based exclusively on the literature, is therefore restricted to the Leucothoe species of the somewhat better known Atlantic Ocean, although also in this area, there is no doubt still a considerable number of as yet undescribed species to be discovered (cf. Thomas and Klebba 2007).
As the genus Leucothoe shows both sexual differences and in many species a considerable allometry, the present key is only suitable for adult male specimens.
Recent authors, for example Thomas and Klebba 2007, have discovered a series of new and most interesting character traits, of use in the identification of Leucothoe species, such as the exact form of the mid–ventral keel and the size and placement of the various rows of setae on the Gn2 propodus. We have unfortunately been unable in most cases to use these characters, as they were not shown in earlier illustrations of other species.
The taxa in the spinicarpa group of species are only now being properly diagnosed and their differences catalogued, and as yet there are, for example, no modern descriptions of L. articulosa (Montagu, 1804) and of L. miersi Stebbing, 1888 (by most authors still synonymized with L. spinicarpa). In addition, we have included both L. occidentalis Reid, 1951 and L. serraticarpa Della Valle, 1893 twice in the key, as the two modern descriptions of these taxa differ in some respect from each other.
We hope that the present key may be a contribution to the further unravelling of the relationships among Atlantic Leucothoe species.
Identification key to Atlantic Leucothoe species (adult males only)
NB We have omitted those taxa considered by White (2011) in her monograph as unidentifiable or ‘probably not valid’. Leucothoe campi Mateus & Mateus, 1986 and L. spinulosa Chevreux, 1919–20, though probably valid species, are not well enough described to be included in the key. All Atlantic records of L. furina (Savigny, 1816) are suspect (cf White 2011), while White’s Atlantic report of L. acanthopus Schellenberg, 1928 rests on a geographic error: the Dahlak Archipelago is in the Red Sea.










Abbreviations
- A 1, 2:
-
Antenna 1, 2
- Acc:
-
Accessory
- Art:
-
Article
- Cx 1–7:
-
Coxa 1–7
- Ep 1–3:
-
Epimera or epimeral plates
- Flag:
-
Flagellum
- Gn 1, 2:
-
Gnathopod 1, 2
- IP:
-
Inner plate
- Md:
-
Mandible
- Mx 1, 2:
-
Maxilla 1, 2
- Mxp:
-
Maxilliped
- OP:
-
Outer plate
- P 3–7:
-
Peraeopod 3–7
- Ped:
-
Peduncle
- Pl 1–3:
-
Pleonite 1–3
- Pn:
-
Peraeonite
- T:
-
Telson
- U 1–3:
-
Uropod 1–3
- Us 1–3:
-
Urosomites 1–3
- Tooth:
-
Non-articulated pointed ectodermal structure
- Spine:
-
Stout, articulated structure (synonymous to “robust seta”)
- Seta:
-
Slender, flexible articulated structure
References
Abildgaard PC (1789) Zoologia Danica seu animalium Daniae et Norvegiae rariorum ac minus notorum. Descriptiones et Historia. Havniae, N. Möller et filius
Bate CS (1862) Catalogue of the specimens of Amphipodous Crustacea in the collections of the British Museum. Taylor & Francis, London
Bate CS, Westwood JO (1863) History of the British sessile-eyed Crustacea. John van Voorst, London
Bruzelius RM (1859) Bidrag til kännedomen om Skandinaviens Amphipoda Gammaridea. Kongl Svensk Vetensk-Akad Handl New Series 3:1–104
Crowe SE (2006) A redescription of Leucothoe spinicarpa (Abildgaard, 1789) based on material from the North Atlantic (Amphipoda, Leucothoidae). Zootaxa 1170:57–68
Dana JD (1852) Conspectus crustaceorum quae in orbis terrarum circumnavigatione, Carolo Wilkes e classe Reipublicae Faederate Duce, lexit et descripsit Jacobus D. Dana. Pars III (Amphipoda 1). Proc Am Acad Arts Sci 2:201–220
Della Valle A (1893) Gammarini del Golfo di Napoli. Fauna Flora Golf Neapel 20:1–948
Gurjanova E (1951) Amphipoda Gammaridea from the seas of the USSR and vicinity. Opredeliteli Faune SSSR Zool Inst Akad Nauk SSSR 41:1–1029 (In Russian)
Krapp-Schickel G (1989) Family Leucothoidae. In: Ruffo S, et al. (eds) The Amphipoda of the Mediterranean. Part 2 Gammaridea (Haustoriidae to Lysianassidae). Mém Inst Océan Monaco 13, pp 443–459
Krapp-Schickel T (2008) What has happened with the Maera- clade (Crustacea, Amphipoda) during the last decades? Boll Mus Civ Stor Natur Verona Botanica Zoologia: 3–32
Krapp-Schickel T, Jarrett NE (2000) The amphipod family Melitidae on the Pacific coast of North America. Part II. The Maera-Ceradocus complex. Amphipacifica 2(4):23–61
Krapp-Schickel T, Menioui M (2005) Leucothoe species from Moroccan Atlantic coasts with redefinition of some species within the Leucothoe spinicarpa clade. Boll Mus Civ Stor Natur Verona Botanica-Zoologia 29:63–83
Lincoln RJ (1979) British marine Amphipoda: Gammaridea. British Museum, London
Mateus A, Mateus E (1986) Campagne de la ‘Calypso’ dans le Golfe de Guinée et aux Iles Principe, São Tomé et Annobon (1956). Amphipodes récoltés à bord de la’Calypso’. Anais Fac Ciénc Porto 66:125–223
Montagu G (1804) Description of several marine animals found on the south coast of Devonshire. Trans Linn Soc London 7:61–85
Norman AM (1869) Shetland final dredging report—Part II. On the Crustacea, Tunicata, Polyzoa, Echinodermata, Actinozoa, Hydrozoa & Porifera. Rep thirty-eighth Meeting Brit Ass Adv Sci 38:113–141
Reid DM (1951) Report on the Amphipoda (Gammaridea and Caprellidea) of the coast of tropical West Africa. Atlantide Rep 2:189–291
Sars GO (1885) Zoology. Crustacea I. Norweg N-Atl Exped 1876–1878 6:1–280
Sars GO (1890–95) Amphipoda. An account of the Crustacea of Norway with short descriptions and figures of all the species I. Alb. Cammermeyer, Christiania & Copenhagen
Schellenberg A (1942) Krebstiere oder Crustacea IV. Flohkrebse oder Amphipoda. Die Tierwelt Deutschlands 40:1–252
Shoemaker CR (1930) The Amphipoda of the Cheticamp expedition of 1917. Contr Can Biol Fish 5:221–359
Stebbing TRR (1888) Report on the Amphipoda collected by H.M.S. Challenger during the years 1873–1876. Rep Sci ResVoyage H.M.S. Challenger 1873–76. Zool 29:1–1737
Stebbing TRR (1906) Amphipoda I. Gammaridea. Das Tierreich 21:1–806
Stephensen K (1923) Crustacea Malacostraca. V. (Amphipoda I). Dan Ingolf-Exped 3:1–100
Stephensen K (1925) Crustacea Malacostraca. VI. (Amphipoda II). Dan Ingolf-Exped 3:101–178
Stephensen K (1931) Crustacea Malacostraca. VII. (Amphipoda III). Dan Ingolf-Exped 3:179–290
Stephensen K (1940) The Amphipoda of N Norway and Spitzbergen with adjacent waters. Tromsø Mus Skr 3(1):279–362
Stephensen K (1944) Crustacea Malacostraca. VIII. (Amphipoda IV). Dan Ingolf-Exped 3(13):1–51
Thomas JD, Klebba KN (2007) New species and host associations of commensal leucothoid amphipods from coral reefs in Florida and Belize. Zootaxa 1494:1–44
von der Brüggen E (1907) Amphipoda. Zoologische Ergebnisse der russischen Expeditionen nach Spitzbergen. Ann Mus Zool Acad Imp Sci St Petersbourg 11:214–244
White KN (2011) A taxonomic review of the Leucothoidae (Crustacea: Amphipoda). Zootaxa 3078:1–113
White KN, Reimer JD (2012a) Commensal Leucothoidae (Crustacea, Amphipoda) of the Ryukyu Archipelago, Japan. Part I: ascidian dwellers. ZooKeys 163:13–55
White KN, Reimer JD (2012b) Commensal Leucothoidae (Crustacea, Amphipoda) of the Ryukyu Archipelago, Japan. Part II: sponge-dwellers. ZooKeys 166:1–58
White KN, Reimer JD (2012c) Commensal Leucothoidae (Crustacea, Amphipoda) of the Ryukyu Archipelago, Japan. Part III. coral-rubble. ZooKeys 173:11–50
Acknowledgments
This study was financially supported by the Norwegian Academy of Sciences, for which we are very grateful. We are also much indebted to the intrepid collectors of Norwegian Sea material, Torleiv Brattegard (Bergen), the late Torleif Holthe and Jon Arne Sneli (both Trondheim), and to the participants of the Skibotn workshop, who have made the sorting of the many samples such a pleasant occasion. Dirk Platvoet (Amsterdam/Leiden) provided again the SEM pictures. Finally, we are very grateful to the reviewers, K. White and O. Coleman, for their valuable comments.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by H.-D. Franke.
Rights and permissions
About this article
Cite this article
Krapp-Schickel, T., Vader, W. Leucothoid and Maerid Amphipods (Crustacea) from deep regions of the North Atlantic. Helgol Mar Res 67, 383–396 (2013). https://doi.org/10.1007/s10152-012-0330-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10152-012-0330-3