- Original Article
- Open access
- Published:
Seven new species of sponges (Porifera) from deep-sea coral mounds at Campos Basin (SW Atlantic)
Helgoland Marine Research volume 70, Article number: 10 (2016)
Abstract
Deep-sea reefs and coral banks are increasingly known as highly biodiverse ecosystems where sponges constitute a significant proportion of builders and inhabitants. Albeit smaller in dimensions, Campos Basin coral mounds also harbor a rich associated fauna, whence only 16 species of sponges had been fully identified this far. Seven new species are described here, viz. Geodia garoupa sp. nov., Vulcanella stylifera sp. nov., Trachyteleia australis sp. nov., Echinostylinos brasiliensis sp. nov., Xestospongia kapne sp. nov., Sympagella tabachnicki sp. nov., and Leucopsacus barracuda sp. nov. Of the 24 species of sponges known from the area, only seven were found elsewhere too, thus suggesting a possible high endemism in Campos Basin. Nevertheless, the widespread occurrence of deep reef-framework building corals along a large sector of the Brazilian coast suggests these habitats and their associated fauna may be more widespread than currently appreciated. Echinostylinos patriciae nom. nov. is proposed for the New Zealand record of E. reticulatus.
Background
Knowledge of the deep sea sponges occurring off Brazil was gained over two important periods, firstly from the H.M.S. Challenger expedition of 1873–1876, and secondly, from a still ongoing effort, that started over 100 years after the first, when a series of, mostly improvised, research ships went off for the first dredgings planned under Project REVIZEE in 1997. This project organized several oceanographic expeditions until 2002, to be followed by Petrobras’ own efforts through several environmental assessment projects, namely OCEANPROF, CAP-BC, ECOPROF and HABITATS. The latter have focused on Campos Basin, Brazil’s main oil producing grounds, and were a response to the country’s environmental authorities’ request for good quality baseline data to support any future need for the evaluation of environmental impacts in Campos Basin. Similar efforts are expanding now to the north (e.g. Espírito Santo and Potiguar Basins) and south (Santos Basin). Preliminary results on the sponges present in these deep sea collections were published by Hajdu [27], Hajdu and Lopes [28], Hajdu et al. [30], Lavrado and Ignacio [42], Lopes and Hajdu [57, 58], Lopes et al. [59–61], Menshenina et al. [64], Muricy et al. [65, 66], Oliveira and Hajdu [68], Rodriguez and Muricy [78], Tabachnick et al. [90], Vieira et al. [107].
The present study reports seven new species collected in the deep waters of Campos Basin (off southeastern Brazil), including the first record of Trachyteleia [96], Echinostylinos [95], and Leucopsacus [35] for the South Atlantic Ocean.
Methods
Campos Basin covers more than 100,000 km2 between the Vitória High (20.5°S) and the Cabo Frio High (24°S) on the Brazilian continental margin. Over 70 % of it lies in depths >200 m [13], and over 85 % of Brazilian crude oil and gas originates from this region. In 2003 PETROBRAS initiated a series of research projects for assessing environmental baseline data. The materials studied here were collected by box-corers, trawls and ROVs, between 744 and 1931 m depth, and are part of the outcome of three umbrella research projects coordinated by CENPES/PETROBRAS: Campos Basin Deep-sea Environmental Project (OCEANPROF), Campos Basin Deep-sea Coral Assessment Project (CAP BC), and Campos Basin Environmental Heterogeneity (HABITATS).
Specimens were identified through the preparation and analysis of dissociated spicules and thick-section mounts, which followed procedures described in [31] for Demospongiae, and [37], for Hexactinellida. The scanning electron microscopes (SEMs) used were a JEOL JSM-6460 LV and a ZEISS DSM-940A from CENPES/PETROBRAS, and a JEOL-6390 LV from Departamento de Invertebrados of Museu Nacional/UFRJ. The newly generated micrometric data for the calculation of means, unless stated otherwise, are derived from 25 spicules of each category for the hexactinellids, and 20 for demosponges, followed by an exhaustive search for maximum and minimum values of length and thickness. The descriptions generated were compared with a tabulation of micrometric, as well as geographic and bathymetric distribution data for all known species in the genera considered, except for Geodia garoupa sp. nov., compared only to species from the South Atlantic, the South Eastern Pacific and the Antarctic. Taxonomic authorities are listed in the comparative tables next to the species considered. Accordingly, they are not referred to in the text.
Following is a list of the abbreviations used and their meanings: CAP BC—Campos Basin Deep-sea Coral Assessment Project; CENPES—PETROBRAS’ Research and Development Center, Rio de Janeiro, RJ, Brazil; HABITATS—Campos Basin Environmental Heterogeneity; MNRJ—Porifera Collection, Museu Nacional, UFRJ, Rio de Janeiro, RJ, Brazil; OCEANPROF—Campos Basin Deep-sea Environmental Project; PETROBRAS—Petróleo Brasileiro S.A., Brazil; UFRJ—Universidade Federal do Rio de Janeiro, Rio de Janeiro, RJ, Brazil.
Results
-
SYSTEMATICS
-
Class DEMOSPONGIAE Sollas, 1885
-
Order TETRACTINELLIDA Marshall, 1876
-
Suborder ASTROPHORINA Sollas, 1887
-
Family GEODIIDAE Gray, 1867
-
Subfamily Geodiinae Sollas, 1888
-
Genus Geodia Lamarck, 1815
-
G. garoupa sp. nov.
-
Fig. 1 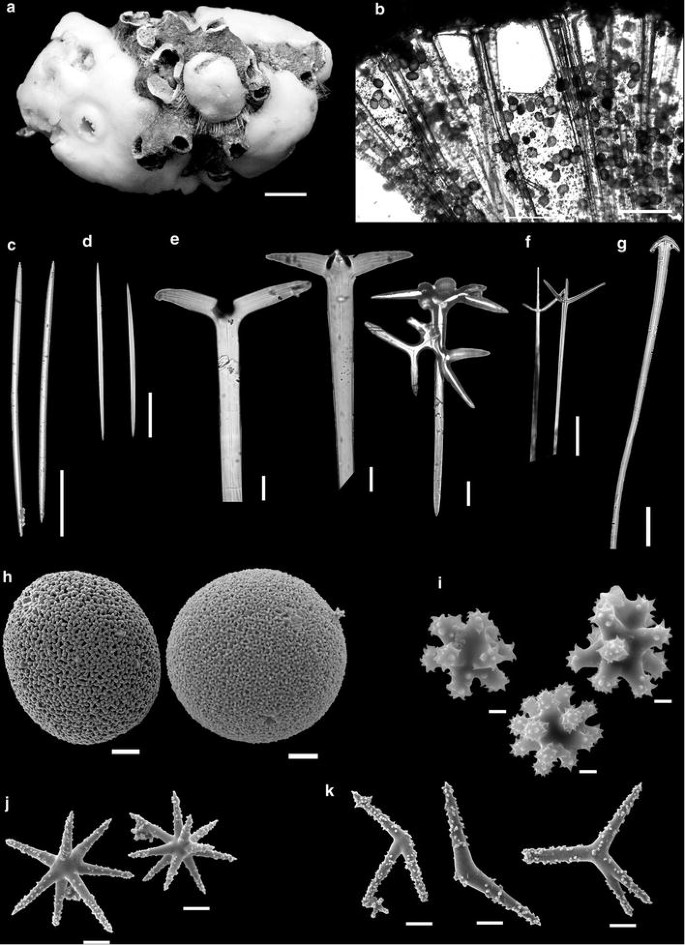
Geodia garoupa sp. nov. a Holotype (MNRJ 7349). b Skeleton. c Oxeas I. d Oxeas II. e Ortho/Dichotriaenes. f Plagiotriaenes. g Anatriaene. h–k SEM of the spicules: h Sterrasters. i Strongylasters. j Oxyasters I. k Oxyasters II. Scales a = 10 mm; b, c = 500 μm; d–g = 100 μm; h = 20 μm; i = 2 μm; j, k = 10 μm
Table 1 Spicule measurements for G. garoupa sp. nov. Minimum − mean − maximum length/width, in μm Table 2 Comparative table of spicular micrometries for the species of Geodia of the South Atlantic Ocean, Magellan Region and Antarctic

Type material
Holotype: OCEANPROF 1, BC-SUL. MNRJ 7349, stn. 4 (Campos Basin, RJ, 22.366°S–39.893°W), 1130 m depth; coll. R/V ‘Astro Garoupa’, demersal fisheries net, 07.ii.2003.
Paratypes: OCEANPROF 1, BC-SUL. MNRJ 7348, 7355, stn. 4 (Campos Basin, RJ, 22.366°S–39.893°W), 1130 m depth; coll. R/V ‘Astro Garoupa’, demersal fisheries net, 07.ii.2003. HABITATS 1. MNRJ 14077, trawl 1 (Campos Basin, RJ, start 23.050°S–41.851°W, end 23.775°S–40.981°W), 1931 m depth; coll. R/V ‘Gyre’, demersal fisheries net, 03.iv.2008. MNRJ 14082, trawl 62 (Campos Basin, RJ), 1244.4 m depth; coll. R/V ‘Gyre’, demersal fisheries net, 29/iv/2008.
Diagnosis
Only Geodia in the Southwest Atlantic and Antarctica with two categories of oxeas, a single category of orthotriaenes tending to dichotriaenes (with or without bifurcated cladomes in different stages of development), plagiotriaenes, and sterrasters always larger than 100 µm.
Description
Habit irregularly massive, cushion-shaped, globular. The holotype (Fig. 1a) is the largest specimen, 7.4 cm in maximum diameter and 4.2 cm thick. Surface hispid, with spicules protruding 1–3 mm in some areas, and possibly removed from most of the surface by damage during trawling. Simple oscules with ~0.5 mm in diameter, pores scattered throughout the surface. Color alive is beige (MNRJ 14082). Consistency hard, but compressible.
Skeleton
Ectosomal skeleton, 1000–1750 µm thick, with small oxeas in the outer surface and a thin layer of small strongylasters, followed by a thick layer of sterrasters supported by the cladomes of the triaenes. The subectosomal region, underneath the cladomes, has large oval canals 308–1525 µm in diameter, with large oxyasters around. Choanosomal skeleton with dense, ascending, and multispicular tracts of oxeas and triaenes (Fig. 1b). Oxyasters and sterrasters are scattered in the skeleton.
Spicules (Table 1)
Oxeas I (Fig. 1c), large, similar to the small ones, tapering gradually, with 1014–3000 µm in length and 50–75 µm thick. Oxeas II (Fig. 1d), small, straight, occasionally curved, tapering abruptly, with 195–227 µm in length and 10–12 µm thick. Orthotriaene (Fig. 1e), with a large size range, small cladomes, with the tips bent down, usually modified to dichotriaenes, with vestigial deuterocladi, 150–300 × 50–100 µm; long, cylindrical rhabdomes, with tips gradually tapering, 1250–3275 µm in length and 50–275 µm thick. Plagiotriaenes (Fig. 1f), very rare and thin, almost always broken, and thus difficult to measure correctly. Anatriaenes (Fig. 1g), found only in the holotype (MNRJ 7349, n = 3), and two paratypes (MNRJ 14077, 14383; n = 1), also very rare and always broken; rhabdome 14.5–25 µm thick, cladi 29–50 µm, and cladome 68–85 µm. Sterrasters (Fig. 1h), spherical to oval, with rays finished by rosettes formed by 4–8 cylindrical actines, 123–144 µm in diameter. Strongylasters (Fig. 1i), small, with 6–12 short and thick rays, strongly spined mainly at the extremity, enlarged centrum, 6–15 µm in diameter. Oxyasters I (to sphaeroxyasters) (Fig. 1j), 8–14 slender and microspined rays, 15–70 µm in diameter. Oxyasters II (Fig. 1k), irregular, with rounded tips, 2–6 microspined rays, 22–90 µm in diameter, very rare, although more abundant in the holotype.
Habitat and distribution
The preserved specimen is attached to a fragment of the coral Solenosmilia variabilis Duncan, 1873. Provisionally endemic from the bathyal zone (1130–1931 m depth) at Campos Basin (SE Brazil).
Etymology
The name garoupa, the Brazilian term for the fish grouper, is used here as a noun in apposition to honor the research vessel, R.V. ‘Astro Garoupa’, on board which OCEANPROF’s materials were collected, the holotype of the new species included.
Remarks
The South Atlantic, South Eastern Pacific and Antarctica harbour 27 species of Geodia (Table 2) according to [105], to which our proposed new species is compared here. Only six of these were reported to bear dichotriaenes, namely australis, libera, magellani, perarmata, robusta, and stellata. Geodia australis is the only of these with oxeas comparably long and thick, but differs from the new species by its possession of protriaenes, as well as very large sterrasters 266–352 µm in diameter. The remaining five species all have considerably larger oxeas than observed in the new species, with the exception of libera, which, instead, has much thinner oxeas. Several other distinctive traits permit further differentiation among these species, a notorious characteristic of species of Geodia and their unusually rich set of morphologic characters. However, one species stands out for its lack of sufficient comparative data to allow a clearcut diagnosis, namely G. cf. reniformis [92] (sensu [98]). No megasclere was associated to this species, thus demanding analysis of its microscleres. The reported two categories of oxyasters, one of spherasters and one of strongylasters (chiasters), differ substantially from the two oxyasters and one strongylaster found in the new species. In addition, while both categories of oxyasters in the new species customarily reach over 50–60 µm, in the Namibian species the smallest category is smaller than 30 µm.
-
Family VULCANELLIDAE Cárdenas, Xavier, Reveillaud, Schander & Rapp, 2011
-
Genus Vulcanella Sollas, 1886
-
Vulcanella stylifera sp. nov.
-
Fig. 2 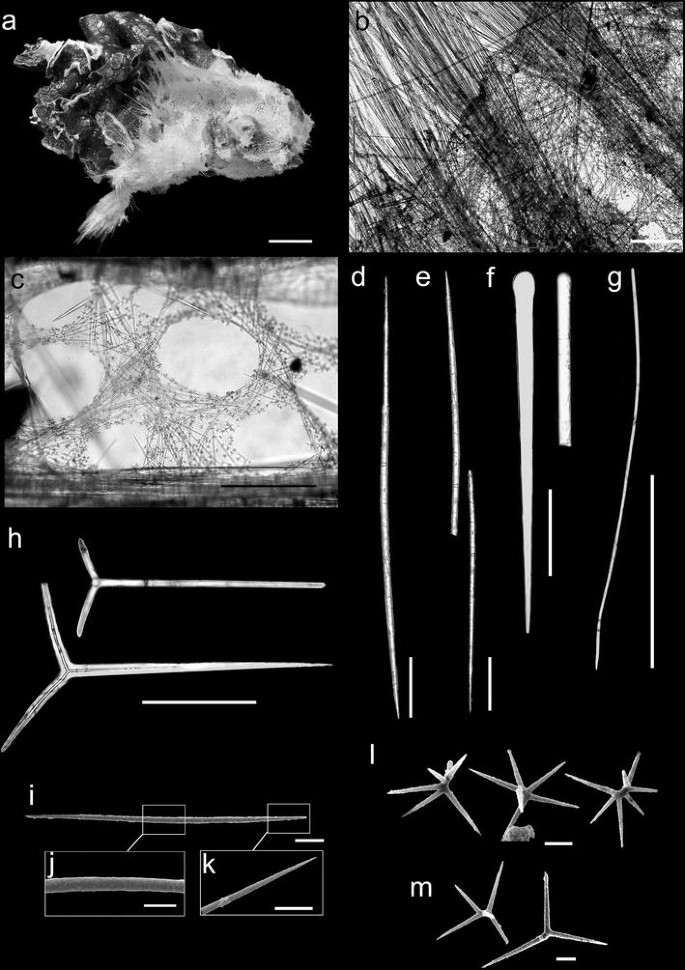
Vulcanella stylifera sp. nov. a Holotype (MNRJ 7343). b Ectosomal skeleton. c Choanosomal skeleton showing the detail of the openings of the aquiferous system. d Oxea I. e Oxea II. f Exotyle. g Style. h Triaenes. i–m SEM of the spicules: i Microxea. j Detail of the middle of the microxea. k Detail of the extremity of the microxea. l Spirasters. m Metasters. Scales a = 1 mm; b, c = 500 μm; d–h = 500 μm; i = 20 μm; j–m = 10 μm
Table 3 Spicule measurements of V. stylifera sp. nov. minimum − mean − maximum length/width, in μm Table 4 Comparative table of spicular micrometries, distribution and depth for the species of Vulcanella

Type material
Holotype: OCEANPROF 1, BC-SUL. MNRJ 7343, stn. 4 (Campos Basin, RJ, start 22.407°S–39.921°W–end 22.366°S–39.893°W), 1128–1135 m depth; coll. R/V ‘Astro Garoupa’, demersal fisheries net, 07.ii.2003.
Paratypes: OCEANPROF 1, BC-SUL. MNRJ 7344, 7345, stn. 4 (Campos Basin, RJ, start 22.407°S–39.921°W–end 22.366°S–39.893°W), 1128–1135 m depth; coll. R/V ‘Astro Garoupa’, demersal fisheries net, 07.ii.2003. MNRJ 7997; stn. 4-2 (Campos Basin, RJ, start 22.407°S–39.926°W–end 22.357°S–39.893°W), 1100 m depth; coll. R/V ‘Astro Garoupa’, demersal fisheries net, 29.viii.2003. OCEANPROF 2, BC-NORTE. MNRJ 7988, stn. 16 (Campos Basin, RJ, start 22.272°S–39.889°W–end 22.219°S–39.870°W), 1059 m depth; coll. R/V ‘Astro Garoupa’, demersal fisheries net, 22.viii.2003.
Diagnosis
Vulcanella with large exotyles (1350–4000 µm long) and styles. Furthermore, no other species in the genus has a combination of oxeas always larger than 2000 µm, triaenes bearing rhabdomes frequently larger than 1000 µm, annulated microxeas frequently over 300 µm long, metasters >40 µm in diameter, and spirasters >30 µm in diameter.
Description
Encrusting or cushion-shaped (Fig. 2a), 66 × 44 × 42 mm (holotype; largest diameter × smaller diameter × thickness) − 21 × 11 × 3 mm (paratype, 7997). Surface hirsute, with a neat sieve-like tangential layer ornamented by large surrounding bundles of megascleres, mainly around the oscules (2–3 mm in diameter), which are irregularly distributed along the surface. Pores were not observed. Consistency is compressible and the specimen is easily broken apart. Colour in ethanol is dark beige to light brown.
Skeleton
Ectosome (Fig. 2b) with bases of triaenes and exotyles, together with dense and almost erect tracts of large oxeas perpendicular to the surface, and piercing it up to 1 cm. It is possible to see some smaller megascleres between these tracts, some of them undoubtedly foreign. Choanosome (Fig. 2c) with a dense, radial architecture of multispicular tracts of oxeas and styles, as well as spirasters and metasters scattered throughout. Annulated oxeas tend to be concentrated around the openings of the aquiferous system, with regular diameter (300–500 µm) and uniform distribution.
Spicules (Table 3)
Oxeas I (Fig. 2d), robust, slightly curved, with extremities abruptly tapered, occasionally strongyloid, 3167–4646 µm long and 38–50 µm thick. Oxeas II (Fig. 2e), echinating, slender, with extremities abruptly tapered, occasionally strongyloid, 5750–12,000 µm long and 20–38 µm thick. Exotyles (Fig. 2f), occasionally strongyloid, robust, straight or slightly curved on the median region, 1350–4000 µm long and 60–75 µm thick. Styles (Fig. 2g), rare, slender, tapering gradually, and slightly curved or flexuous, 1050–2525 µm long and 5–27.5 µm thick. Orthotriaenes (Fig. 2h), robust, ranging to plagiotriaenes; terminations usually thin, only occasionally strongyloid; rhabdomes, 875–1725 µm long and 50–75 µm thick; cladomes, 525–1200 µm long and 40–50 µm thick. Annulated microxeas (Fig. 2i–k), straight or slightly curved, tapering gradually, rings clearly visible at the central region, occasionally centrotylote, 130–446 µm long and 1.8–2.5 µm thick. Spirasters (Fig. 2l), 6–8 rays, with two or more twists, 28–40 µm in largest diameter. Metasters (Fig. 2m), 4–6 rays, 43–48 µm in largest diameter.
Habitat and distribution
Provisionally endemic from the bathyal zone of Campos Basin (SE Brazil), 1059–1130 m depth, where the species appears to be moderately common, occurring associated with S. variabilis.
Etymology
The specific epithet stylifera is derived from the species possession of styles.
Remarks
The exotyles seen in the new species might be triaenes with reduced cladomes, as apparent from their slightly conical shape, similar to rhabdomes in general. However, due to the lack of intermediate forms that might support the hypothesis of suppression of the cladomes, we prefer to believe these spicules to be truly monaxonic. In addition, it is not possible to see any bi/trifurcation at the basal extremity of the axial filament, which could indicate the spicule’s poliaxonic character.
Vulcanella has recently been allocated in Vulcanellidae [12] based on molecular data. These authors also raised subgenus Annulastrella [62] to full generic status, and transferred it to Theneidae. Triaenes have been secondarily lost in this genus. As a consequence, there no longer is an undisputed subgeneric classification for Vulcanella.
Fourteen species of Vulcanella are known worldwide (Table 4): Vulcanella aberrans, V. acanthoxea, V. bifacialis, V. cribrifera, V. cribriporosa, V. doederleini, V. gracilis, V. horrida, V. netheides, V. orthotriaena, V. osculanigera, V. porosa, V. theneides and V. tricornis. None of these possess a category of styles, but malformed triaenes may be common (e.g. [62]). Nevertheless, to the best of our knowledge, the clearly recognizable category seen in the new species has not been reported previously in the literature, and appears to us a convincing support for recognition of the new species. Rather than relying onthis single diagnostic character, a thorough comparison was made where every known species was contrasted to the newly proposed one, and every spicule category considered.
The echinating oxeas in the new species can frequently be 10,000 µm long, but they will never be as large as those in Vulcanella tricornis, which attains 25,000 µm. On the other hand, several species reach only up to 5000–6000 µm, and can be confidently distinguished on the basis of this character alone. These species are V. aberrans, V. cribriporosa, V. doederleini, V. gracilis, V. orthotriaena and V. osculanigera. Smaller rhabdomes in the triaenes also distinguish a series of species: V. aberrans, V. bifacialis, V. cribrifera, V. cribriporosa, V. netheides, V. orthotriaena, V. porosa and V. theneides. Microxeas that are consistently smaller than those of the new species occur in V. acanthoxea, V. cribrifera, V. cribriporosa and V. doederleini (with, in addition, a second category of much smaller microxeas). Metasters and/or spirasters can also be consistently smaller, as is the case of those in V. aberrans, V. acanthoxea, V. bifacialis, V. cribrifera, V. cribriporosa, V. gracilis, V. horrida, V. netheides, V. porosa and V. tricornis. It is obvious from the above remarks that the new species has spicules which frequently reach quite larger dimensions when contrasted to the same categories in other Vulcanella spp. Some species possess calthrops, a spicule category absent from the new species. This character adds further support for the distinction of V. acanthoxea, V. cribrifera, V. cribriporosa, V. doederleini, V. gracilis, V. horrida, V. osculanigera and V. tricornis.
The genus was already registered for the South Atlantic [30], although this is the first full species description for a Vulcanella from this area.
-
Order POLYMASTIIDA Morrow & Cárdenas, 2015
-
Family POLYMASTIIDAE Gray, 1867
-
Genus Trachyteleia Topsent, 1928
-
Trachyteleia australis sp. nov.
-
Fig. 3 
Trachyteleia australis sp. nov. a Holotype (MNRJ 7359). b Choanosomal skeleton. c Tylostyles I. d Tylostyles II. e Exotyles. f, g SEM of the spicules: f base of the exotyle; g detail of the spines of exotyle. h Subtylostyles. Scales a = 5 mm; b = 500 μm; c–e, h = 100 μm; f, g = 10 μm
Table 5 Spicule measurements of Trachyteleia australis sp. nov. minimum – mean − maximum length/width, in μm Table 6 Comparative table of spicule micrometries, distribution and depth for the species of Trachyteleia

Type material
Holotype: OCEANPROF 1, BC-NORTE. MNRJ 7359, stn. 18 (Campos Basin, RJ, start 22.270°S–39.791°W–end 22.221°S–39.789°W), 1622–1628 m depth, coll. R/V ‘Astro Garoupa’, demersal fisheries net, 12.ii.2003.
Paratypes: OCEANPROF 1, BC-NORTE. MNRJ 7358, stn. 18 (Campos Basin, RJ, start 22.270°S–39.791°W–end 22.221°S–39.789°W), 1622–1628 m depth, coll. R/V ‘Astro Garoupa’, demersal fisheries net, 12.ii.2003. OCEANPROF 2, BC-SUL. MNRJ 8014, stn. 2-1 (Campos Basin, RJ, start 22.512°S–40.016°W–end 22.479°S–39.977°W), 1107–1141 m depth, coll. R/V ‘Astro Garoupa’, demersal fisheries net, 27.viii.2003.
Diagnosis
Trachyteleia with ectosomal tylostyles up to 500 µm long and 22 µm thick.
Description
Massive fragments with 1 × 1 cm. Surface hispid with echinating spicules (Fig. 3a). Consistency hard, only slightly compressible. Color alive is beige to yellow, and light to dark-beige after fixation with ethanol.
Skeleton
Ectosomal skeleton a dense layer of tylostyles, traversed by echinating choanosomal subtylostyles and exotyles. Choanosomal skeleton radial, with dense, multispicular ascending tracts of choanosomal tylostyles, exotyles and subtylostyles (Fig. 3b).
Spicules (Table 5)
Tylostyles I (Fig. 3c), ectosomal, smooth, straight, markedly fusiform, robust, with well developed tyle, 155–504 µm long and 9–22 µm thick. Tylostyles II (Fig. 3d), choanosomal, smooth, slightly curved at the base, slightly fusiform, relatively slender, 271–747 µm long and 6–22 µm thick. Exotyles (Fig. 3e–g), choanosomal, rough, or microspined in the central region, 611–1484 µm long and 7–17 µm thick. Choanosomal subtylostyles (Fig. 3h), large and smooth, 659–3841 µm long and 8–31 µm thick.
Habitat and distribution
Provisionally endemic from the bathyal zone of Campos Basin (SE Brazil), at 1107–1628 m depth.
Etymology
The specific epithet, australis, is the Latin word for “from the South”, which highlights the fact that this is the first record of Trachyteleia for the Southern Hemisphere.
Remarks
Only two species were previously known in Trachyteleia, T. hispida and T. stephensi, recognized by the presence of distally microspined exotyles [8] similar to those found in the Brazilian specimens.
Nevertheless, several differences separate the new species from the other two previously known. The ectosomal tylostyles I in T. australis sp. nov. are larger than those in T. hispida (up to 180 µm) and in T. stephensi (up to 275 µm). Intermediate and principal tylostyles are not always well separated in T. hispida and T. stephensi sensu [96], as well as in the new species. In T. australis sp. nov. they are larger than the intermediate/principal tylostyles of T. stephensi, and smaller than the same category in T. hispida (Table 6). Exotyles are much smaller in T. stephensi than in the new species (up to 735 and 2377 µm, respectively). While in T. hispida they can be about 2600 µm long, similar to the larger ones in T. australis sp. nov.
This is the first record of the genus for the South Atlantic, and the whole Southern Hemisphere.
-
Order Poecilosclerida Topsent, 1928
-
Family Phellodermidae van Soest & Hajdu, 2002
-
Genus Echinostylinos Topsent, 1927
-
Echinostylinos brasiliensis sp. nov.
-
Fig. 4 
Echinostylinos brasiliensis sp. nov. a Holotype (MNRJ 8003c). b Skeleton. c Styles. d SEM of the isochelae. e Isochelae. Scales a = 10 mm, b = 500 μm, c = 100 μm, d = 5 μm, e = 10 μm
Table 7 Spicule micrometries for E. brasiliensis sp. nov. minimum − mean − maximum length/width, in μm Table 8 Comparative table of spicule micrometries, distribution and depth for the species of Echinostylinos

Type material
Holotype: OCEANPROF 1, BC-SUL, MNRJ 7328, stn. 4 (Campos Basin, RJ, start 22.407°S–39.921°W–end 22.366°S–39.893°W), 1130 m depth; coll. R/V ‘Astro Garoupa’, demersal fisheries net, 07.ii.2003.
Paratype: OCEANPROF 2, BC-SUL, MNRJ 8001c, MNRJ 8003c, stn. 4-2 (Campos Basin, RJ, start 22.407°S–39.926°W–end 22.357°S–39.893°W), 1100 m depth; coll. R/V ‘Astro Garoupa’, demersal fisheries net, 29.viii.2003.
Diagnosis
The only species of Echinostylinos with a single category of megascleres, and markedly curved unguiferate isochelae 22–30 µm long.
Description
The holotype and largest specimen is encrusting an area 2.3 × 1.7 cm, with short, irregular, anastomosing branches (Fig. 4a). Its surface is slightly hispid, and no special ornamentation, pores, or oscules have been observed. Consistency is compressible and color in ethanol beige.
Skeleton
Choanosomal skeleton organized, with tracts of styles, occasionally reticulated, or some styles crossing the tracts (Fig. 4b). Spongin can be present in several points, and isochelae are abundant all over the sponge.
Spicules (Table 7)
Megascleres (Fig. 4c)—styles varying little in dimensions, smooth, slightly curved, tapering gradually to sharp ends, 776–1067 µm long and 17.5–22.5 µm thick. Microscleres (Fig. 4d, e)—unguiferate isochelae with a markedly curved shaft in profile view, and four sharp alae on each extremity, 22–30 µm.
Habitat and distribution
Provisionally endemic from the bathyal zone of Campos Basin (SE Brazil), at 1100–1130 m depth.
Etymology
The specific epithet, brasiliensis, relates to its type locality, off south-eastern Brazil, and for this being the sole known species of the genus reported from the Brazilian coast.
Remarks
Ten species have been allocated in Echinostylinos, nine of which remain accepted as valid: E. glomeris, E. gorgonopsis, E. hirsutus, E. mycaloides, E. reticulatus, E. schmidtii, E. shimushirensis, E. stylophora and E. tubiformis. Echinostylinos unguiferus has been transferred to Monanchora on account of the anchorate nature of its isochelae (referred to as unguiferate by Esteves [25], Van Soest [102]). On the basis of the remarks offered by Van Soest and Hajdu [104] on the unlikelihood of the conspecificity of New Zealand and Azorean records of E. reticulatus, we decided to propose a new name for the former record, namely E. patriciae nom.nov. (honouring Dame Patricia Bergquist, first author of the record—holotype NIWA 105240, paratpe NIWA 105473). This is done not just because of the distant localities of occurrence and remarkably distinct bathymetry of both, but also to highlight some morphological divergence such as the much thicker megascleres and smaller sigmas in the type specimen of E. reticulatus.
Table 8 summarizes the morphological data and known distribution of every species, and was used for a detailed comparison with the new species. The genus was formerly known from the North Atlantic, NW and Center-South Pacific, from a depth range of 55–2500 m. The new species described is the first record of Echinostylinos for the entire South Atlantic, and is clearly distinguished from the remaining species in having only a single category of megascleres, as well as unguiferate isochelae, shorter than 30 μm.
The styles of E. brasiliensis sp. nov. are smaller than those of E. glomeris, E. hirsutus, E. shimushirensis, and E. tubiformis; larger than those of E. gorgonopsis, E. reticulatus, E. patriciae nom.nov., E. schmidti, and E. stylophora; and similar to those of E. mycaloides.
Further, the new species from Campos Basin has isochelae that are smaller than those of E. glomeris, E. gorgonopsis, E. mycaloides, E. patriciae nom.nov., E. reticulatus, E. shimushirensis, E. schmidtii, E. stylophora, and E. tubiformis; and similar to those of E. hirsutus, although this species has two categories of megascleres. Considering the shape of the isochelae, E. mycaloides is the species coming closer to E. brasiliensis sp. nov., as both share the unguiferate, strongly arched pattern in their isochelae. The former species has, nevertheless, isochelae 3× as large as those in the new one. Other species have notoriously distinct isochelae morphologies, frequently much stouter, with spatulate alae. An exception is E. hirsutus, of dubious affinities, with much reduced isochelae of unclear morphology. This species should be reexamined under SEM for a sounder assignment to Echinostylinos.
Echinostylinos schmidtii has been only poorly illustrated by Arnesen [2] and needs to be redescribed. Unfortunately the whereabouts of its type material could not be traced in Bergen or Oslo and it appears to be lost (H.T. Rapp, pers. comm. on 2014 09 26). In principle it has a single category of styles, which would bring it close to the new species, but the large sigmas and the seemingly typical arcuate spatulate isochelae would set Arnesen’s species far from the Brazilian material described above.
Echinostylinos shimushirensis and E. tubiformis might be synonymous, as both share a similar spicule set, with quite comparable spicule dimensions (Table 8). Both species were also recorded from the same depth zone, but given their far apart occurrence (7600 km away), and slightly divergent habit and isochelae morphology, we prefer to hold their formal synonimization until the type materials of both have been reexamined.
-
Order Haplosclerida Topsent, 1928
-
Family Petrosiidae van Soest, 1980
-
Genus Xestospongia de Laubenfels, 1932
-
Xestospongia kapne sp. nov.
-
Fig. 5 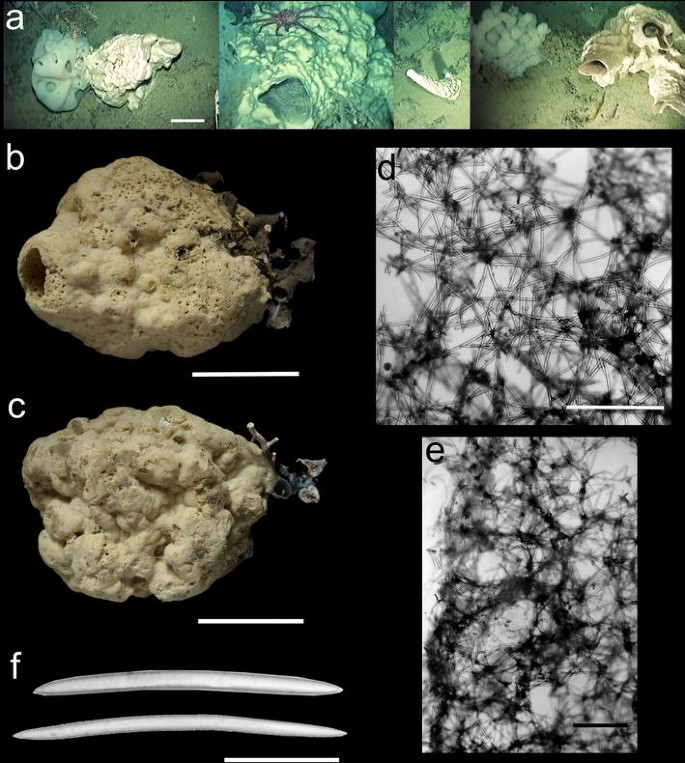
Xestospongia kapne sp. nov. a Several specimens in situ. b, c Holotype preserved in ethanol. d Ectosomal skeleton. e Choanosomal skeleton. f Oxeas. Scales a–c = 10 cm; d, e = 500 μm; f = 100 μm
Table 9 Comparative table of spicule micrometries, distribution and depth data for the species of Xestospongia de Laubenfels, 1932

Type material
Holotype: CAP BC, MNRJ 13541, Caratinga Oil Field 9.5″, bank 3 (Campos Basin, RJ, 22.623°S–40.264°W), 923 m depth, coll. R/V ‘Toisa Conqueror’, ROV, 12/iii/2006.
Comparative material Petrosia friabilis Topsent, 1892 (holotype, MOM 04 0159)
Diagnosis
Xestospongia with oxeas ranging from 200 to 300 µm in length, barrel- or chimney-shaped, found at deep waters (877–1053 m).
Description
Massive, robust, barrel- or chimney-shaped, with an irregular contour and a large apical pseudo-oscule (Fig. 5a–c). The surface has lobular projections and shallow depressions or gaps, and rough texture. Several specimens were recognized from video footage, where it was nicknamed “cannon-sponge”. Some of these form clusters of up to three chimneys, and the largest specimens recorded were presumably nearly 50 cm tall.
Skeleton
Ectosomal skeleton with a loose reticulation formed by the tangential arrangement of spicules composing the terminal tufts of ascending choanosomal spiculo-fibers (Fig. 5d). Ectosomal meshes are seen here and there, with 140–250 µm in diameter. Choanosomal architecture with a reticulation that is clearly visible only in some areas, overlaid by abundant oxeas in confusion (Fig. 5e). Meshes are only seldom observed, being 200–400 µm in diameter. The distinction between primary ascending fibers and interconnecting secondaries is unclear.
Spicules
Oxeas (Fig. 5f)—Relatively robust, usually slightly curved, tapering gradually, with 204–265.1–301 µm in length and 11–12.6–14 µm thick. Derived and rare forms have more accentuated curvatures, and sometimes styloid or subtylostyloid ends.
Habitat and distribution
Provisionally endemic from the bathyal zone of Campos Basin (SE Brazil), at 877–1053 m depth.
Etymology
The specific epithet relates to the chimney-like habit of the species (kapne = Greek for chimney), and is used here as a noun in apposition.
Remarks
The genus Xestospongia is found from the intertidal to 1800 m depth. It has ca. 35 species known worldwide, 12 of which occurring in the Atlantic Ocean [105]. Only X. grayi and X. muta were known from Brazil until now, both from warm and shallow waters of the Northeastern coast, and they differ from the new species due to the presence of strongyles as megascleres [32, 67]. Likewise, another 13 species also possess strongyles in their spicule set (see Table 9), thus differing from the new species. The possession of sigmas sets X. bocatorensis, X. edapha, and X. emphasis, apart from X. kapne sp. nov.
Only seven species of Xestospongia were known to occur deeper than 100 m (Table 9): X. clavata, X. coralloides, X. diprosopia, X. friabilis, X. hispida, sensu [98], X. rampa, and X. variabilis, sensu [93]. All of them possess oxeas as megascleres, except for X. rampa, which has strongyles and can be easily differentiated from the new species through this character. X. clavata, X. diprosopia, and X. variabilis, sensu [93] have oxeas that are always larger than 300 µm long and 30 µm thick, being thus also considerably distinct from the new species. Despite the possession of oxeas smaller than 300 µm in length, X. coralloides, X. friabilis and X. hispida, sensu [98] are quite distinct from the new species too. The oxeas in X. friabilis can be much smaller than those in X. kapne sp. nov., with a minimum length of 116 µm (holotype remeasured; Table 9), in contrast to a minimum of 204 µm in the new species. X. hispida, sensu [98], from Namibia, has oxeas whose minimum length is much longer than observed in the smallest oxeas of the new species (290 × 204 µm, respectively). The habit of X. coralloides and X. hispida is also a distinctive character, being flat-lammelate and lobate, respectively, with no tubular structures topped by large pseudoscules.
Some Xestospongia spp. approach the new species in having only oxeas and in a similar size range (Table 9): X. bergquistia, X. delaubenfelsi, X. mammillata, X. novaezelandiae, X. plana, X. tuberosa, and X. vansoesti [4]. In addition to most of these species being shallow water dwellers (down to 55 m depth), further slight differences appear when contrasting them to X. kapne sp. nov. X. bergquistia was described as an erect, cup-shaped sponge, thus reminiscent of the barrel-shaped new species. Nevertheless, the former species possibly has two categories of oxeas, one of them much thinner (2–8 µm thick), and the other with occasional styloid ends, which coupled to its very far distribution and shallow-water habit, renders the hypothesis of co-especificity with the new species highly unlikely. Very thin oxeas are also found in X. mammillata and X. vansoesti (1.5 and 2 µm thick, the smallest ones, respectively), although no suggestion has been made of these being anything but young forms. Xestospongia delaubenfelsi and X. plana have a flattened habit too, which is considerably distinct from the barrel-shape of the new species. Besides, they are known only from very far, biogeographically unrelated localities (Chile and the Mediterranean, respectively), and in none of them oxeas reach 300 µm in length. Xestospongia novaezelandiae (from New Zealand) is erect massive and irregular, and X. tuberosa (from the Western Indian Oc.) is lobate, thus also differentiating in shape from the new species.
-
Class Hexactinellida Schmidt, 1870
-
Subclass Hexasterophora Schulze, 1886
-
Order Lyssacinosida Zittel, 1877
-
Family Rossellidae Schulze, 1885
-
Genus Sympagella Schmidt, 1870
-
Sympagella tabachnicki sp. nov.
-
Fig. 6 
Sympagella tabachnicki sp. nov. a Holotype (MNRJ 13365), a1 detail of the dermal surface and a2 base. b–h SEM of the spicules: b ends of the choanosomal diactins, c hypoatrial pentactin, d dermal pinular, e, f atrial pinular hexactins, g discohexaster, and h onychexaster. Holotype: d, e. Paratype: b, c, f–h (MNRJ 7315A). Scales: a = 50 mm; b = 20 μm; c = 100 μm; d–f = 50 μm; g, h = 10 μm
Fig. 7 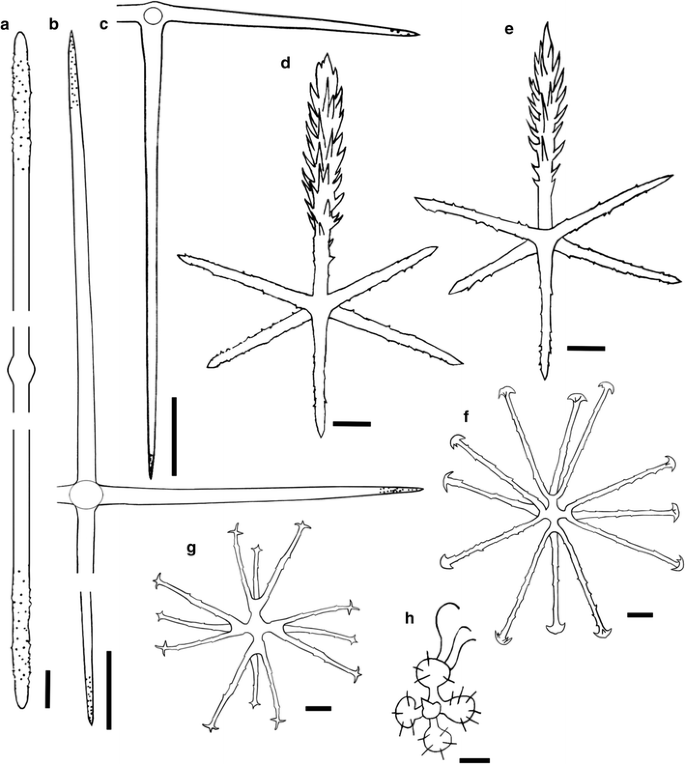
Sympagella tabachnicki sp. nov. Spicules. a Choanosomal diactin, b choanosomal hexactin, c hypoatrial pentactin, d dermal pinular hexactin, e atrial pinular hexactin, f discohexaster, g onychexaster, and h strobiloplumicome. Holotype: a, c–g. Paratypes: b (MNRJ 13362) and h (MNRJ 7315A). Scales a, d, e = 20 μm; b, c = 100 μm; f, g = 10 μm; h = 5 μm
Table 10 Spicule micrometries for S. tabachnicki sp. nov Table 11 Comparative morphological and distributional data for the species of Sympagella Schmidt, 1870


Type material
Holotype: CAP BC Rota Gas, MNRJ 13365, trecho 1, Espadarte Field (Campos Basin, RJ, 22.757°S–40.433°W), 945 m depth, coll. R/V ‘Toisa Conqueror’, ROV, 22/iii/2005.
Paratypes: OCEANPROF 1, BC-SUL, MNRJ 7315A, 7316, stn 4 (Campos Basin, RJ, 22.366°S–39.893°W), 1128–1135 m depth, coll. R/V ‘Astro Garoupa’, demersal fisheries net, 07/ii/2003. CAP BC Duto Gas, MNRJ 13362, trecho 1, Espadarte Field (Campos Basin, RJ, 22.757°S–40.433°W), 945 m deep, coll. R/V ‘Toisa Conqueror’, ROV, 22/iii/2005.
Comparative material: Sympagella nux Schmidt, 1870—USNM 7588 (holotype, one dissociated spicule slide).
Diagnosis
Sympagella without prostalia lateralia, composed by diactins and hexactins as single choanosomal spicules; dermal and atrial pinular hexactins with columnar-shaped pinular rays; hypodermal and hypoatrial pentactins present; strobiloplumicomes, discohexasters and onychexasters as microscleres.
Description
Basiphytose sponge with saccular body, thin walls (ca. 1 mm), attached to solid substrate by a base (Fig. 6a). Holotype is 115 long × 154 wide × 16 mm thick. Paratypes are composed of fragments.
Skeleton
Choanosomal skeleton composed of diactins and rare hexactins. Hypodermalia and hypoatrialia are pentactins. Dermalia and atrialia are pinular hexactins. Basalia are hexactins fused to each other by synapticules.
Spicules (Table 10)
Choanosomal diactins are smooth and curved, with conical and microspined ends (Figs. 6b, 7a). Choanosomal hexactins have slightly curved rays with microspined ends (at least in the distal and proximal rays), which gradually taper (Fig. 7a). Hypodermalia and hypoatrialia are smooth pentactins with microspined and pointed ends (Figs. 6c, 7c). Dermalia and atrialia are hexactins with variably developed pinular rays bearing short spines; tangential and proximal rays are microspined, with conical or pointed ends (Figs. 6d–f, 7d, e). Microscleres are strobiloplumicomes, discohexasters and onychexasteres. Typical strobiloplumicomes are always found broken (Fig. 7h). Discohexasters (Figs. 6g, 7f) are spherical with toothed discs, with short primary rays and three long, spined, secondary rays. Onychexasters (Figs. 6h, 7g) have short primary rays and three long, microspined, secondary rays. In addition, a series of rare microscleres was observed, but judged to be of external origin: oxyhexasters (68–113 µm, n = 17), discohexactins (75–115, n = 4), onychexactins (58–60 µm, n = 2) and oxyhexactins (55–145 µm, n = 12) were found in the paratypes; and rare hemioxyhexasters (95–130 µm, n = 3), in the holotype and paratypes as well.
Habitat and distribution
This species is known only from its type locality in the SW Atlantic (Espadarte Field, Campos Basin, SE Brazil), 945–1135 m.
Etymology
The proposed name, tabachnicki, is in honour of Dr. Konstantin Tabachnick (Institute of Oceanology, Russian Academy of Science), who already described over 120 hexactinellid taxa from all around the world, including the Campos Basin area.
Remarks
The genus presently contains nine recognized species distributed in the Atlantic and Pacific Oceans, and Antarctica, between 27 and 4063 m depth (Table 11): S. anomala from NO Pacific, S. cantharellus from Central-NO Pacific, S. clavipinula from New Caledonia, S. cooki and S. ecomari from the Northern Mid-Atlantic Ridge, S. gracilis from Indonesia, S. johnstoni from Antarctica and Subantarctic Islands, S. multihexastera from NW Australia, and S. nux from the Mediterranean, Cape Verde Islands, the Caribbean, and SE Atlantic.
The Japanese S. anomala has been recently re-examined by [37], who have emphasized the pinular rays of its hexactins to be lanceolate in shape (thickest in the middle, fusiform). From Ijima’s original description [36] it is apparent that only dermalia are lanceolate. Pinular hexactins of the Brazilian species have columnar-shaped pinular rays. Furthermore, Ijima’s species was found to bear diactins as large as 2000 µm only, in great contrast to the 6000 µm reached by a few spicules in the new species. Exceptionally, S. anomala may bear choanosomal tauactins and stauractins, and its hypodermal/hypoatrial pentactins can be absent, which further distinguish the Japanese and Brazilian species.
Lèvi and Lèvi [53] described a rather deviating specimen of S. anomala from the Philippines, albeit judging it to be decidedly conspecific to the type. This specimen has a much larger and stouter category of choanosomal diactins (up to 6000/70–80 µm), as well as oxyhexasters. These authors did not mention the presence of choanosomal hexactins, tauactins and stauractins. We find the cospecificity hypothesis to be very unlikely, but the clarification of this issue is beyond the scope of the present contribution. Instead, we must figure how the Philippines’ material differs from the new species presently described. This is obvious in the shape of the dermal pinules, which are lanceolated in the Philippines’ sponge, the extremely stout choanosomal diactins (up to 70/80 µm), and the oxyhexasters, which are lacking in the new species.
The new species differs from S. cantharellus by the possession of choanosomal diactins considerably smaller and thinner (1075–6200/8–25 vs. 2200–9100/5–80 µm), which can be spined in the Pacific species. Additionally, S. cantharellus has a mushroom-like body form and oxyhexasters, whereas the new species has a saccular body form and no oxy-tipped microscleres. The hypodermal/hypoatrial pentactins can be absent in S. cantharellus.
Important distinguishing features for the new species in comparison to S. clavipinula are the presence of hypodermal/hypoatrial pentactins with smaller tangential rays (210–550/15–30 in the former vs. 340–1000/23–46 µm in the latter) and absence of oxyhexasters (in the former). Additionally, S. clavipinula has dermal pinular hexactins with spherical, club-shaped pinular rays (artichoke-like). Rare choanosomal stauractins were found in the New Caledonian species, whereas we failed to find a single spicule in the four S. Atlantic specimens described here.
Sympagella tabachnicki sp. nov. differs from N Atlantic Sympagella species by the absence of hypodermal/hypoatrial hexactins and the presence of onychexasters. The only species in the genus with diactins as the sole choanosomal megasclere are S. anomala, S. cooki and S. ecomari, which sets them apart from the new species who has choanosomal hexactins in addition. Furthermore, S. cooki possesses oxy-tipped microscleres (oxyhexasters, hemioxyhexasters and oxyhexactins), absent in the S. Atlantic species. Also, rare pinular pentactins are found in both species from N. Atlantic, but not in the new species.
The new species can be differentiated from S. gracilis by the latter’s much thicker choanosomal diactins (60–100 µm), dermal pinular hexactins with relatively smaller pinular rays (100–150/20–30 µm), possession of oxyhexasters, and lack of discohexasters.
Sympagella johnstoni is the only species in the genus with prostalia lateralia and conules in the body wall. The new species further differs from S. johnstoni by the latter’s much larger choanosomal diactins (1800–10,000 µm) and possession of hypodermal/hypoatrial hexactins and discohexactins. Janussen et al. [37] reported diactins in S. johnstoni to be up to 500 µm thick, but this is a likely mistake, as the figure provided by these authors illustrates a 50 µm thick diactin, much more in accordance to values observed in other species of Sympagella, albeit still much thicker than the new species.
The new species differs from S. multihexastera by the latter’s much smaller and thicker choanosomal diactins (1500–2900/10–130 µm), choanosomal hexactins with smaller rays (150–370 µm), dermal and atrial pinular hexactins with relatively smaller tangential and proximal rays (dermal: 41–77 µm; atrial: 41–71, 51–82 µm) and possession of oxyoidal microscleres (oxyhexasters, hemioxyhexasters and oxyhexactins).
Sympagella tabachnicki sp. nov. differs from S. nux by the latter’s presence of dermal pinular pentactins (instead of hexactins), tylohexasters, oxyhexasters, oxyhexactins and microdiasters, and absence of onychexasters. In addition, S. nux has much larger and thinner atrial pinular hexactins (76–836/4–11 µm), and rare hypodermal/hypoatrial stauractins.
The Japanese S. anomala and S. johnstoni can be considered closer to new species by the presence of choanosomal diactins and hexactins, hypodermal/hypoatrial pentactins, pinular hexactins and only disco and onycho-tipped microscleres. However, the new species described here is distinguished from all species of Sympagella by one or more characters quoted above.
-
Family Leucopsacidae Ijima, 1903
-
Genus Leucopsacus Ijima, 1898
-
Leucopsacus barracuda sp. nov.
-
Fig. 8 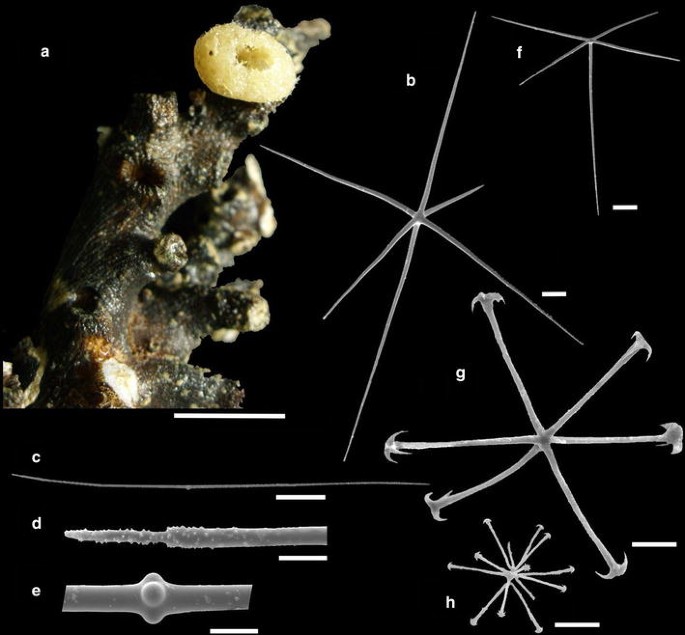
Leucopsacus barracuda sp. nov. a Holotype (MNRJ 13368). b–h SEM of the spicules: b choanosomal hexactin, c choanosomal diactin, d end of the choanosomal diactin, e central knob of the choanosomal diactin, f dermal pentactin, g anchorate discohexactin, and h discohexaster. Holotype: b–g. Paratype: h (MNRJ 13369). Scales a = 10 mm; b, f = 100 μm; c = 200 μm; d, e, g, h = 20 μm
Table 12 Spicule measurements of L. barracuda sp. nov Table 13 Comparative morphological and distributional data for the species of Leucopsacus Ijima, 1898

Type material
Holotype: CAP BC Barracuda, MNRJ 13368, area 40 line 37 (Campos Basin, RJ, 22.522°S–40.246°W), 744 m deep, coll. R/V ‘Toisa Conqueror’, ROV, 17/iii/2006.
Paratype: CAP BC Caratinga Oil Field 9.5’, MNRJ 13369, bank 3 (Campos Basin, RJ, 22.623°S–40.264°W), 923 m deep, coll. R/V ‘Toisa Conqueror’, ROV, 12/iii/2006.
Diagnosis
Leucopsacus with diactins and hexactins as choanosomal spicules, dermal pentactins and small discohexasters (up to 80 μm) as microscleres, in addition to anchorate discohexactins and rare hemidiscohexasters.
Description
Basiphytose sponge with ovoid body, thin walls (ca. 2 mm), attached to solid substrate (coral) by a base (Fig. 8a). Holotype is 8 mm long × 7 mm wide, with a short peduncle about 1 mm in length, and osculum 3 mm in diameter. Paratype is composed by a fragment, 7 mm long × 10 mm wide × 1 mm thick.
Skeleton
Choanosomal skeleton composed by hexactins and diactins. Dermalia are pentactins and basalia are hexactines fused to each other by synapticules.
Spicules (Table 12)
Choanosomal hexactins are smooth, with straight or curved rays and pointed ends (Fig. 8b). Choanosomal diactins are smooth and curved, with a central knob and microspined pointy ends (Fig. 8c–e). Dermalia are smooth pentactins with conical and microspined ends (Fig. 8f). Microscleres are anchorate discohexactins and discohexasters. One hemidiscohexaster was found in the holotype and it was considered proper. Anchorate discohexactins have long and microspined rays with toothed discs (Fig. 8g). Discohexasters (Fig. 8h) have short primary rays and four long, microspined secondary rays.
Habitat and distribution
This species is known only from its type locality in the SW Atlantic (Barracuda Field, Campos Basin, SE Brazil), 744–923 m. Holotype was collected epibiontic on dead Enallopsammia rostrata (Pourtalès, 1878).
Etimology
The proposed name, barracuda, is a noun in apposition, derived from the species’ type locality, Barracuda Oil Field (Campos Basin, SW Atlantic).
Remarks
The genus presently contains four recognized species distributed in the Atlantic and Pacific Oceans, between 214 and 1378 m depth (Table 13): L. distantus from New Caledonia, L. ingolfi from Boreal Atlantic, L. orthodocus from Japan, and L. scoliodocus from Japan and Cape Verde Island. Leucopsacus ingolfi is closest to the new species by the presence of choanosomal diactins and absence of atrial hexactins. However, the new species differs from N Atlantic species by the presence of larger choanosomal hexactins (380–1334 vs. 300 μm), dermal pentactins (tangencial rays 287–677 and proximal rays 493–1047 vs. tangencial rays 180 and proximal rays 400 μm) and anchorate discohexactins (95–165 vs. 70 μm) and, much smaller discohexasters (38–78 vs. 440 μm).
Discussion
Deep sea sponge grounds are considered Vulnerable Marine Ecosystems (VME; [34]). This is so because (1) they are important as shelter and nursery for juvenile life-stages, (2) are fragile (low recovery prognosis), (3) usually harbor long-lived and slow-growing dominant species, (4) and can increase habitat heterogeneity (ecological processes dependent on the highly structured system). Despite the fact that, given data available now, there seems to be no deep-sea sponge reefs nor ostur (Geodia banks) in the Campos Basin area, there occur nevertheless important sponge aggregations associated to the deep-sea coral mounds [56, 58, 61], and these are obviously fragile and vulnerable.
For some time, notable deep-sea coral mounds were known to occur at Campos Basin. Viana et al. [106] reported these to extend for hundreds of meters in length and tens in width, reaching up to 15 m in height, and spreading over at least 40 km. Similar structures were later reported from the perimeter of seabed pockmarks at Santos Basin (700 m depth) by Sumida et al. [84], who interpreted their finding as supporting evidence for the widespread occurrence of coral banks off SE Brazil. In both cases, it is the highly oxygenated and vigorously flowing Antarctic Intermediate Waters, as well as likely seepage of light hydrocarbons that seem to render possible the occurrence of these complex communities. The uniqueness along the Brazilian coast of these SE Brazilian deep sea habitats is unlikely. This is so in view of the occurrence of isolated records of azooxanthelate reef-building corals off most of the Brazilian coast [14, 39, 69], and of similar prevailing environmental conditions along a vast stretch of the NE South-American slope (Brazil’s SE and NE slope) [15, 17, 56]. Lopes et al. [59] drew attention to the fact that Brazil was not equipped for state-of-the-art academic studies of deep sea habitats then, due to lack of appropriate oceanographic vessels. This scenario has been changing lately, so that great expectations exist on forthcoming findings of deep-sea coral and sponge banks on the Brazilian continental slope.
References
Arnesen E. Spongien von der norwegischen Küste. ii, Monaxonida: Halichondrina. Bergens Museum Åarbog. 1903;1–30, pls I–VII.
Baer L. Silicispongien von Sansibar, Kapstadt and Papeete (inaugural dissertation: Berlin). Arch Naturgesch. 1906;72(1, 1):1–32, pls I–V.
Bakus GJ, Nishiyama GK. Three species of toxic sponges from Cebu, Philippines (Porifera: Demospongiae). Proc Biol Soc Wash. 2000;113(4):1162–72.
Bergquist PR, Fromont PJ. The Marine Fauna of New Zealand: Porifera, Demospongiae, Part 4 (Poecilosclerida). NZ Oceanogr Inst Mem. 1988;96:1–197.
Bergquist PR, Warne KP. The Marine Fauna of New Zealand: Porifera, Demospongiae, Part 3 (Haplosclerida and Nepheliospongida). NZ Oceanogr Inst Mem. 1980;87:1–77.
Boury-Esnault N. Family Polymastiidae Gray, 1867. In: Hooper JNA, Van Soest RWM, editors. Systema Porifera. A guide to the classification of sponges. New York: Kluwer; 2002. p. 201–19.
Burton M. Description of South African sponges collected in the South African Marine Survey. Part I. Myxospongia and Astrotetraxonida. Fisheries bulletin. Fisheries and Marine Biological Survey Division, Union of South Africa Reports No. 4 (special rept. 9); 1926. p. 1–29, 6 pls.
Burton M. Hexactinellida. Danish Ingolf Exp. 1928;6(4):1–18.
Burton M. Sponges. In: Fridriksson A, Tuxen SL, editors. The Zoology of Iceland, vol 2. Ejnar Munksgaard: Copenhagen& Reykjavik; 1959. p. 1–71.
Cárdenas P, Rapp HT. A review of Norwegian streptaster-bearing Astrophorida (Porifera: Demospongiae: Tetractinellida), new records and a new species. Zootaxa. 2012;3253:1–52.
Cárdenas P, Xavier JR, Reveillaud J, Schander C, Rapp HT. Molecular phylogeny of the Astrophorida (Porifera, Demospongiae) reveals an unexpected high level of spicule homoplasy. PLoS One. 2011;6(4):e18318. doi:10.1371/journal.pone.0018318.
Carminatti M, Scarton JC. Sequence stratigraphy of the Oligocene turbidite complex of the Campos Basin, Offshore Brazil: an overview. In: Weimer P, Link MH, editors. Seismic facies and sedimentary processes of submarine fans and turbidite systems. New York: Springer; 1991. p. 41–246.
Cordeiro RTS, Kitahara MV, Amaral FD. New records and range extensions of azooxanthellate scleractinians (Cnidaria: Anthozoa) from Brazil. Mar Biodivers Rec. 2012;5:1–6.
Coutinho PN. Oceanografia Geológica. In: Levantamento do Estado da Arte da Pesquisa dos Recursos Vivos Marinhos do Brasil–Programa REVIZEE. Relatório Consolidado. Rio de Janeiro, Brasilia: FEMAR/SECIRM/MMA; 1996. Downloaded from http://www.mma.gov.br/estruturas/revizee/_arquivos/levarte.pdf on 02 Nov 2014.
da Silva CMM, Mothes B. Three new species of Geodia Lamarck, 1815 (Porifera, Demospongiae) from the bathyal depths off Brazilian coast, Southwestern Atlantic. Rev Suisse Zool. 2000;107(1):31–48.
Damuth JE, Hayes DE. Echo character of the East Brazilian Continental Margin and its relationship to sedimentary processes. Mar Geol. 1977;24:73–95.
de Laubenfels MW. The marine fauna and fresh-water sponges of California. Proc US Natl Mus. 1932;81(4):1–140.
de Laubenfels MW. New sponges from the Puerto Rican deep. Smiths Misc Coll. 1934;91(17):1–28.
de Laubenfels MW. The sponges of California. In: Abstracts of dissertations for the degree of doctor of philosophy, Stanford University Bulletin No. (5), vol. 5; 1930. p. 24–9.
de Laubenfels MW. The sponges of the West-Central Pacific. Oregon State monographs. Stud Zool. 1954;7:i–x, 1–306, pls I–XII.
de Laubenfels MW. The sponges of woods hole and adjacent waters. Bull Mus Comp Zool. 1949;103(1):1–55, pls 1–3.
Dendy A. Porifera. Part I: non-Antarctic sponges, British Antarctic (“Terra Nova”) expedition, 1910. Nat Hist Rep Zool. 1924;6(3):269–392.
Díaz MC, Thacker RW, Rützler K, Piantoni C. Two new haplosclerid sponges from Caribbean Panama with symbiotic filamentous cyanobacteria, and an overview of sponge-cyanobacteria associations. In: Custódio MR, Lôbo-Hajdu G, Hajdu E, Muricy G, editors. Porifera research—biodiversity, innovation and sustainability, vol. 28. Rio de Janeiro: Museu Nacional, Série Livros; 2007. p. 31–9.
Dickinson MG. Sponges of the Gulf of California. Allan Hancock Pac Exped. 1945;11(1):1–252.
Esteves EL. Revisão Taxonômica e Filogenia de Crambe Vosmaer, 1880 e Monanchora Carter, 1883 (Crambeidae, Poecilosclerida, Demospongiae). Ph.D. thesis, Museu Nacional—Universidade Federal do Rio de Janeiro. 2009.
Fromont J. Descriptions of species of the Petrosida (Porifera:Demospongiae) occurring in the tropical waters of the Great Barrier Reef. Beagle Rec N Terr Mus Arts Sci. 1991;8:73–96.
Hajdu E. Porifera. In: Lavrado HP, Brasil ACS, editors. Biodiversidade da Região Oceânica Profunda da Bacia de Campos: Megafauna e Ictiofauna Demersal. Rio de Janeiro: SAG Serv; 2010. p. 31–79.
Hajdu E, Lopes DA. Checklist of Brazilian deep-sea sponges. In: Custódio MR, Lôbo-Hajdu G, Hajdu E, Muricy G, editors. Porifera research—biodiversity, innovation and sustainability, vol. 28. Rio de Janeiro: Museu Nacional, Série Livros; 2007. p. 353–9.
Hajdu E, Muricy G, Custodio M, Russo C, Peixinho S. Geodia corticostylifera (Demospongiae, Porifera) new astrophorid from the Brazilian coast (Southwestern Atlantic). Bull Mar Sci. 1992;51(2):204–17.
Hajdu E, Santos CP, Lopes DA, Oliveira MV, Moreira MCF, Carvalho MS, Klautau M. Filo Porifera. In: Amaral ACZ, Rossi-Wongtschowski CLDB, editors. Biodiversidade Bêntica da região sul-sudeste brasileira. São Paulo: REVIZEE Score Sul—Bentos; 2004. p. 49–56.
Hajdu E, Peixinho S, Fernandez JCC. Esponjas marinhas da Bahia—Guia de campo e laboratório. Rio de Janeiro: Museu Nacional, Série Livros; 2011.
Hechtel GJ. New species of marine Demospongiae from Brazil. Iheringia Sér Zool. 1983;63:58–89.
Hentschel E. Die Kiesel- und Hornschwämme des Nördlichen Eismeers. In: Römer F, Schaudinn F, Brauer A, Arndt W, editors. Fauna Arctica. Eine Zusammenstellung der arktischen Tierformen mit besonderer Berücksichtigung des Spitzbergen-Gebietes auf Grund der Ergebnisse der Deutschen Expedition in das Nördliche Eismeer im Jahre 1898, vol 5; 1929. p. 857–1042, pls XII–XIV (G. Fischer, Jena).
Hogg MM, Tendal OS, Conway KW, Pomponi SA, van Soest RWM, Gutt J, Krautter M, Roberts JM. Deep-sea sponge grounds: reservoirs of biodiversity. UNEP-WCMC Biodiversity Series No. 32. Cambridge: UNEP-WCMC; 2010. http://www.unep-wcmc.org/resources/publications/UNEP_WCMC_bio_series/32.aspx (Accessed 02 Sep 2014).
Ijima I. The genera and species of Rosselidae. Annot Zool Jap. 1898;2(2):41–55.
Ijima I. Studies on the Hexactinellida. Contribution III (Placosoma, a New Euplectellid; Leucopsacidae and Caulophacidae). J Coll Sci Imp Univ Tokyo. 1903;18(1):1–124, pls I–VIII.
Janussen D, Tabachnick KR, Tendal OS. Deep-sea Hexactinellida (Porifera) of the Weddell Sea. Deep Sea Res Part II. 2004;51:1857–992.
Keller C. Die Spongienfauna des Rothen Meeres (II. Halfte). Z Wiss Zool ABT A. 1891;52:294–368, pls XVI–XX.
Kitahara MV. Species richness and distribution of azooxanthellate Scleractinia in Brazilian waters. Bull Mar Sci. 2007;81(3):497–518.
Koltun VM. Sponge fauna of the northwestern Pacific from the shallows to the hadal depths. In: Bogorov VG, editor. Fauna of the Kurile-Kamchatka Trench and its environment, vol 86. Moskwa: Institute of Oceanology of the Academy of Sciences of the U.S.S.R. Akademiya Nauk SSSR. Trudy Instituta Okeanologii in P.P. Shishov and Izdatelstvo Nauka; 1970. p. 165–221.
Koltun VM. Sponges of the Antarctic. 1. Tetraxonida and Cornacuspongida. In: Pavlovskii EP, Andriyashev AP, Ushakov PV, editors. Biological reports of the Soviet Antarctic Expedition (1955–1958). Akademya Nauk SSSR; 1964. p. 6–133, 443–48 (English translation, 1966, Israel Program for Scientific Translation).
Lavrado HP, Ignacio BL. Biodiversidade bentônica da região central da Zona Econômica Exclusiva brasileira, vol. 18. Rio de Janeiro: Museu Nacional, Série Livros; 2006.
Lebwohl F. Japanische Tetraxonida, I. Sigmatophora und II. Astrophora metastrosa. J Coll Sci Imp Univ Tokyo. 1914;35(2):1–116, pls I–IX.
Lehnert H, van Soest RWM. More North Jamaican deep fore-reef sponges. Beaufortia. 1999;49(12):141–69.
Lévi C. Spongiaires de la côte du Sénégal. Bull Inst Fr Afr Noire (A. Sciences naturelles). 1952;14(1):34–59.
Lévi C. Résultats scientifiques des Campagnes de la ‘Calypso’.Campagne 1954 dans l’Océan Indien (suite). 2. Les spongiaires de l’Ile Aldabra. Ann Inst Oceanogr. 1961;39(1):1–32, pls 1–2.
Lévi C. Spongiaires du Vema Seamount (Atlantique Sud). Bull Mus Natl Hist Nat (2). 1969;41(4):952–73.
Lévi C. Porifera Demospongiae: Spongiaires bathyaux de Nouvelle-Calédonie, récoltés par le ‘Jean Charcot’. Campagne BIOCAL, 1985. pp. 9–87. In: Crosnier A, editor. Résultats des campagnes MUSORSTOM, vol 11. Mem Mus Nat Hist Nat Nouv Ser A. 1993;158:1–426.
Lévi C. Spongiaires d’Afrique du Sud. (3) Tetractinellides. Trans R Soc S Afr. 1967;37:227–56, pls XVII–XIX.
Lévi C. Spongiaires des zones bathyale, abyssale et hadale. Galathea Report. In: Scientific results of the Danish deep-sea expedition round the world, 1950–1952. 1964;7:63–112, pls II–XI.
Lévi C, Lévi P. Eponges Tétractinellides et Lithistides bathyales de Nouvelle-Calédonie. Bull Mus Natl Hist Nat (4). 1983;5(1):101–68.
Lévi C, Lévi P. Démosponges bathyales récoltées par le N/O‘Vauban’ au sud de la Nouvelle-Calédonie. Bull Mus Natl Hist Nat (4, A). 1983;5(4):931–97.
Lévi C, Lévi P (1989). Spongiaires (MUSORSTOM 1 and 2). In: Forest J, editor. Résultats des Campagnes MUSORSTOM, vol 4. Mem Mus Nat Hist Nat Nouv Ser A. 1989;143:25–103.
Lévi C, Vacelet J. Éponges récoltées dans l’Atlantique oriental par le ‘Président Théodore-Tissier’ (1955–1956). Rev Trav Inst Pech Marit. 1958;22(2):225–46.
Little FJ Jr. The sponge fauna of the St. George’s Sound, Apalache Bay, and Panama City Regions of the Florida Gulf Coast. Tulane Stud Zool. 1963;11(2):31–71.
Lopes DA. Taxonomia de Cladorhizidae (Poecilosclerida, Demospongiae) do mar profundo chileno e brasileiro, com ênfase em poríferos associados à Scleractinia azooxantelados (Cnidaria). Análise Cladística e Biogeográfica de Asbestopluma (Asbestopluma). Ph.D. thesis, Museu Nacional—Universidade Federal do Rio de Janeiro. 2011.
Lopes DA, Hajdu E. Two new Mycalina from the Se Brazilian shelf and slope collected by Programme REVIZEE (Poecilosclerida, Demospongiae). J Mar Biol Assoc UK. 2004;84:25–8.
Lopes DA, Hajdu E. Carnivorous sponges from deep-sea coral mounds in Campos Basin (SW Atlantic), with description of six new species (Cladorhizidae, Poecilosclerida, Demospongiae). Mar Biol Res. 2014;10:329–56.
Lopes DA, Hajdu E, Reiswig HM. Redescription of two Hexactinosida (Porifera, Hexactinellida) from the southwestern Atlantic, collected by programme REVIZEE. Zootaxa. 2005;1066:43–56.
Lopes DA, Hajdu E, Reiswig HM. Taxonomy of Euretidae (Porifera, Hexactinellida, Hexactinosida) of Campos Basin, southwestern Atlantic, with a description of a new species. Mar Biol Res. 2007;3:243–55.
Lopes DA, Hajdu E, Reiswig HM. Taxonomy of Farrea (Porifera, Hexactinellida, Hexactinosida) from the southwestern Atlantic, with description of a new species and a discussion on the recognition of subspecies in Porifera. Can J Zoolog. 2011;89:169–89.
Maldonado M. Family Pachastrellidae Carter, 1875. In: Hooper JNA, van Soest RWM, editors. Systema Porifera. A guide to the classification of sponges. New York: Kluwer; 2002. p. 141–62.
Maldonado M, Uriz M-J. A new species of Sphinctrella (Demospongiae: Astrophorida) and remarks on the status of the genus in the Mediterranean. Med K Belg Inst Nat Wet. 1996;66(Suppl.):175–84.
Menshenina LL, Tabachnick KR, Janussen D. Revision of the genus Neopsacas (Hexactinellida, Rossellidae, Crateromorpha) with the description of new species and subspecies. Zootaxa. 2007;1463:55–68.
Muricy G, Santos CP, Batista DC, Lopes DA, Pagnoncelli D, Monteiro LC, Oliveira MV, Carvalho M, Melão M, Moreira MCF, Klautau M, Rodriguez PRD, Costa RN, Silvano R. Filo Porifera. In: Lavrado HP, Ignacio BL, editors. Biodiversidade bentônica da região central da Zona Econômica Exclusiva brasileira, vol. 18. Rio de Janeiro: Museu Nacional, Série Livros; 2006. p. 109–45.
Muricy G, Hajdu E, Oliveira MV, Heim AS, Costa RN, Lopes DA, Melão M, Rodriguez PRD, Silvano RG, Monteiro LC, Santos C. Filo Porifera. In: Lavrado HP, Viana MS, editors. Atlas de invertebrados marinhos da região central da Zona Econômica Exclusiva brasileira, vol 25. Rio de Janeiro: Parte 1. Museu Nacional, Série Livros; 2007. p. 25–57.
Muricy G, Esteves EL, Moraes F, Santos JP, Silva SM, Klautau M, Lanna E. Biodiversidade Marinha da Bacia Potiguar—Porifera. Rio de Janeiro: Museu Nacional, Série livros; 2008. p. 29.
Oliveira MV, Hajdu E. Taxonomy of Rhabderemia Topsent, 1890 collected from the south-eastern Brazilian continental shelf and slope by Programme REVIZEE (Rhabderemiidae, Poecilosclerida, Demospongiae), with the description of two new species. Zootaxa. 2005;844:1–12.
Pires DO. The azooxanthellate coral fauna of Brazil. In: George RY, Cairns SD, editors. Conservation and adaptive management of seamount and deep-sea coral ecosystems. Coral Gables: Rosenstiel School of Marine and Atmospheric Science, University of Miami; 2007. p. 265–72.
Pulitzer-Finali G. A collection of marine sponges from East Africa. Ann Mus Civ Stor Nat “Giacomo Doria”. 1993;89:247–350.
Pulitzer-Finali G. Sponges from the Bismarck Sea. Boll Mus Ist Biol Univ Genova. 1996;60–61:101–38.
Pulitzer-Finali G. A collection of West Indian Demospongiae (Porifera). In appendix, a list of the Demospongiae hitherto recorded from the West Indies. Ann Mus Civ Stor Nat “Giacomo Doria”. 1986;86:65–216.
Pulitzer-Finali G. Some new or little-known sponges from the Great Barrier Reef of Australia. Boll Mus Ist Biol Univ Genova. 1980–1982;48–49:87–141.
Ridley SO, Dendy A. Preliminary report on the Monaxonida collected by H.M.S. ‘Challenger’. Ann Mag Nat Hist (5). 1886;18, 325–51, 470–93.
Ridley SO. Spongiida. In: Report on the zoological collections made in the Indo-Pacific Ocean during the Voyage of H.M.S. ‘Alert’, 1881–1882. London: British Museum (Natural History); 1884. p. 366–482, pls 39–43; 582–630, pls 53–54.
Ristau DA. Six new species of shallow-water marine demosponges from California. Proc Biol Soc Wash. 1978;91(3):569–89.
Riveros-Zuñiga F. Xestospongia delaubenfelsi nov.sp. (Haliclonidae-Porifera) (1 fig). Rev Bio Mar. 1951;3(3):226–28.
Rodriguez PRD, Muricy G. A new species of Cinachyra (Demospongiae: Tetillidae) collected by Project REVIZEE off Espírito Santo State, SE Brazil. In: Custódio MR, Lôbo-Hajdu G, Hajdu E, Muricy G, editors. Porifera research: biodiversity, innovation and sustainability, vol. 28. Rio de Janeiro: Museu Nacional, Série Livros; 2007. p. 547–53.
Samaai T, Gibbons MJ. Demospongiae taxonomy and biodiversity of the Benguela region on the west coast of South Africa. Afr Nat Hist. 2005;1:1–96.
Schmidt O. Grundzüge einer Spongien-Fauna des atlantischen Gebietes. Leipzig: Wilhelm Engelmann; 1870. iii–iv, 1–88, pls I–VI.
Schulze FE. Caulophacus arcticus (Armauer Hansen) und Calycosoma gracile F.E. Sch. nov. spec. Abhandlungen der Koeniglichen Akademie der Wissenschaften zu Berlin; 1903. p. 1–22, pls I–II.
Silva CMM. Revisão das espécies de Geodia Lamarck, 1815 (Porifera, Astrophorida, Geodiidae) do Atlântico Ocidental e Pacífico Oriental. Ph.D. thesis, Universidade de São Paulo. 2002.
Sollas WJ. Report on the Tetractinellida collected by H.M.S. Challenger, during the years 1873–1876. Report on the scientific results of the voyage of H.M.S. Challenger, 1873–1876. Zoology. 1888;25(63):1–458, pls I–XLIV.
Sumida PYG, Yoshinaga MY, Madureira LAS, Hovland M. Seabed pockmarks associated with deepwater corals off SE Brazilian continental slope, Santos Basin. Mar Geol. 2004;207(1–4):159–67.
Tabachnick KR. Family Rossellidae Schulze, 1885. In: Hooper JNA, van Soest RWM, editors. Systema Porifera. A guide to the classification of sponges. New York: Kluwer; 2002. p. 1441–505.
Tabachnick KR. Family Leucopsacidae Ijima, 1903. In: Hooper JNA, van Soest RWM, editors. Systema Porifera. A guide to the classification of sponges. New York: Kluwer; 2002. p. 1435–40.
Tabachnick KR, Lévi C. Lyssacinosa du Pacifique sudouest (Porifera: Hexactinellida). In: Marshall B, Richer de Forges B, editors. Tropical deep-sea benthos no. 23. Mem. Mus. Natl. Hist. Nat., vol 191; 2004. p. 11–71.
Tabachnick KR, Menshenina LL. New data on glass sponges (Porifera, Hexactinellida) of the northern Mid-Atlantic Ridge. Part 2. Aphrocallistidae, Euretidae, Euplectellidae and Rossellidae (with descriptions of two new species of Sympagella). Mar Biol Res. 2013;9:469–87.
Tabachnick KR, Janussen D, Menshenina LL. New Australian Hexactinellida (Porifera) with a revision of Euplectella aspergillum. Zootaxa. 2008;1866:7–68.
Tabachnick KR, Menshenina LL, Lopes DA, Hajdu E. Two new Hyalonema species (Hyalonematidae: Amphidiscosida) from eastern and south-eastern Brazil, and futher Hexactinellida (Porifera) collected from seamounts off south-eastern Brazil by the RV ‘Marion Dufresne’ MD55 expedition. J Mar Biol Assoc UK. 2009;89:1243–50.
Tanita S, Hoshino T. The demospongiae of Sagami Bay. Japan: Biological Laboratory, Imperial Household; 1989. i–xiii, 1–197 (in English), pls 1–19; 1–166 (in Japanese).
Thiele J. Studien über pazifische Spongien. I. Japanische Demospongien. Zoologica. Stuttgart: Original-Abhandlungen aus dem Gesamtgebiete der Zoologie. 1898;24(1):1–72, pls I–VIII.
Topsent E. Contribution à l’étude des Spongiaires de l’Atlantique Nord (Golfe de Gascogne, Terre-Neuve, Açores). Rés Camp scient Prince Albert I. Monaco. 1892;2:1–165, pls I–XI.
Topsent E. Notes sur les Eponges du Travailleur et du Talisman. III. Leucopsacus scoliodocus Ij. var. retroscissus; Sarostegia oculata Tops. Bull Mus Hist nat Paris. 1904;10:372–8.
Topsent E. Diagnoses d’Éponges nouvelles recueillies par le Prince Albert ler de Monaco. Bull Inst Océanogr Monaco. 1927;502:1–19.
Topsent E. Spongiaires de l’Atlantique et de la Méditerranée provenant des croisières du Prince Albert ler de Monaco. Rés Camp scient Prince Albert I. Monaco. 1928;74:1–376, pls I–XI.
Topsent E. Contribution nouvelle à la connaissance des Eponges des côtes d’Algérie. Les espèces nouvelles d’O. Schmidt, 1868. Bull Inst Océanogr Monaco. 1938;758:1–32.
Uriz MJ. Deep-water sponges from the continental shelf and slope off Namibia (Southwest Africa): Classses Hexactinellida and Demospongia. Monogr Zool Marina. 1988;3:9–157.
Vacelet J, Vasseur P. Spongiaires des grottes et surplombs des récifs de Tuléar (Madagascar). Recl Trav Stn Mar Endoume. 1965;2–4:71–123.
Vacelet J, Vasseur P, Lévi C. Spongiaires de la pente externe des récifs coralliens de Tuléar (sud-ouest de Madagascar). Mem Mus Nat Hist Nat Ser A Zool. 1976;49:1–116.
Van Soest RWM. Marine sponges from Curaçao and other Caribbean localities. Part II. Haplosclerida. Stud Fauna Curacao Caribb Isl. 1980;62(191):3–132.
Van Soest RWM. Marine sponges from Curaçao and other Caribbean localities. Part III. Poecilosclerida. Stud Fauna Curacao Caribb Isl. 1984;66(199):1–167.
Van Soest RWM, De Weerdt WH. New records of Xestospongia species (Haplosclerida: Petrosiidae) from the Curaçao reefs, with a description of a new species. Beaufortia. 2001;51(7):109–17.
Van Soest RWM, Hajdu E. Family Phellodermidae fam. nov. In: Hooper JNA, VanSoest RWM, editors. Systema Porifera. A guide to the classification of sponges. New York: Kluwer; 2002. p. 621–4.
Van Soest RWM, Boury-Esnault N, Hooper JNA, Rützler K, de Voogd NJ, Alvarez de Glasby B, Hajdu E, Pisera AB, Manconi R, Schoenberg C, Janussen D, Tabachnick KR, Klautau M, Picton B, Kelly M, Vacelet J, Dohrmann M, Díaz MC, Cárdenas P. World Porifera database. 2015. http://www.marinespecies.org/porifera (Accessed July 2015).
Viana AR, Faugères JC, Kowsmann RO, Lima JAM, Caddah LFG, Rizzo JG. Hydrology, morphology and sedimentology of the Campos continental margin, offshore Brazil. Sediment Geol. 1998;115:133–57.
Vieira WF, Cosme B, Hajdu E. Three new species of Erylus (Demospongiae, Astrophorida, Geodiidae) from the Almirante Saldanha Seamount (off SE Brazil), with further data for a tabular review of worldwide species and comments on Brazilian seamount sponges. Mar Biol Res. 2010;6(5):437–60.
von Lendenfeld R. Die Tetraxonia. Wissenschaftliche Ergebnisse der Deutschen Tiefsee-Expedition auf der Dampfer Valdivia 1898–1899. 1907;11(1–2):i–iv, 59–374, pls IX–XLVI.
von Lendenfeld R. The Sponges. 3. Hexactinellida. In: Reports on the scientific results of the expedition to the Eastern Tropical Pacific, in charge of Alexander Agassiz, by the U.S. Fish Commission Steamer ‘Albatross’, from October, 1904, to March, 1905, Lieut. Commander L.M. Garrett, U.S.N., Commanding, and of other expeditions of the ‘Albatross’, 1891–1899 (29). Mem Mus Comp Zool Harvard. 1915;42(2):1–396, pls 1–109.
Wilson HV. Reports on an exploration off the West Coasts of Mexico, Central and South America, and off the Galapagos Islands, in charge of Alexander Agassiz, by the U.S. Fish Commission Steamer ‘Albatross’ during 1891, Lieut. Commander Z.L. Tanner, U.S.S., commanding. XXX. The Sponges. Mem Mus Comp Zool Harvard. 1904;30(1):1–164, pls 1–26.
Wilson HV. Silicious and horny sponges collected by the U.S. Fisheries Steamer ‘Albatross’ during the Philippine Expedition, 1907–1910. In: Contributions to the biology of the Philippine Archipelago and adjacent regions. Bull US Natl Mus. 1925;100(2, part 4):273–532, pls 37–52.
Authors’ contributions
MSC concluded descriptions and plates of Demospongiae, drafted species’ remarks and article discussion. DAL identified and described the Hexactinellida. BC drafted species descriptions and plates for the Demospongiae, aside Xestospongia. EH took part in identification of all species, in rounding up descriptions, remarks, tables, plates and the discussion. All authors read and approved the final manuscript.
Acknowledgements
Prof. Dr. Michelle Kelly and Ms. Diana Macpherson (NIWA—National Institute of Water and Atmospheric Research, New Zealand) are deeply thanked for tracking, finding, registering, and photographing the specimens here designated holotype and paratype of Echinostylinos patriciae nom.nov. The same applies to Ms. Michèle Bruni (MOM—Oceanographic Museum of Monaco, Monaco), who kindly sent on loan the type of Petrosia friabilis Topsent, 1892; Prof. Dr. Allen Collins (National Museum of Natural History, Smithsonian Institution, USA), who granted accesss to the holotype of Sympagella nux Schmidt, 1870; and Prof. Dr. Hans Tore Rapp (University of Bergen, Norway) for his efforts to find the current whereabouts of Echinostylinos schmidti (Arnesen, 1903), alas unsuccessfully. CENPES–PETROBRAS is thanked for the invitation to take part in projects OCEANPROF and CAP BC, as well as for granting access to the institution’s SEM equipment, operated by Rogério da Silva Martins da Costa, Aílton Luiz da Silva de Souza, and Rose Maria de Lima Mencarelli. We are thankful also to Elivaldo de Lima and Amanda da Veiga for SEM operation at the Center for Scanning Electron Microscopy of Museu Nacional/UFRJ, as well as Sula Salani Mota for help with preparations. The establishment of this Center was made possible by a grant from CENPES–PETROBRAS, and is part of the company’s thematic network for marine environmental monitoring. Débora de Oliveira Pires of Museu Nacional is thanked for the identification of scleractinian corals. CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior), CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico), FAPERJ (Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro), and CENPES–PETROBRAS are deeply thanked for the provision of grants and (or) fellowships.
Competing interests
The authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
de S. Carvalho, M., Lopes, D.A., Cosme, B. et al. Seven new species of sponges (Porifera) from deep-sea coral mounds at Campos Basin (SW Atlantic). Helgol Mar Res 70, 10 (2016). https://doi.org/10.1186/s10152-016-0461-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s10152-016-0461-z